
Abstract
Purpose
Common Lymphatic Endothelial and Vascular Endothelial Receptor 1 (Clever-1) is expressed by a subset of immunosuppressive macrophages and targeting the receptor with therapeutic antibodies has been shown to activate T-cell-mediated anti-cancer immunity. The aim of this research was to study Clever-1 expression in breast cancer. Specifically, how Clever-1 + macrophages correlate with clinicopathologic factors, Tumor Infiltrating Lymphocytes (TILs) and prognosis.
Methods
Tissue microarray blocks were made from 373 primary breast cancer operation specimens. Hematoxylin and Eosin (H&E-staining) and immunohistochemical staining with Clever-1, CD3, CD4 and CD8 antibodies were performed. Differences in quantities of Clever-1 + macrophages and TILs were analyzed. Clever-1 + cell numbers were correlated with 25-year follow-up survival data and with breast cancer clinicopathologic parameters.
Results
Low numbers of intratumoral Clever-1 + cells were found to be an independent adverse prognostic sign. Increased numbers of Clever-1 + cells were found in high grade tumors and hormone receptor negative tumors. Tumors that had higher amounts of Clever-1 + cells also tended to have higher amounts of TILs.
Conclusion
The association of intratumoral Clever-1 + macrophages with better prognosis might stem from the function of Clever as a scavenger receptor that modulates tumor stroma. The association of Clever-1 + macrophages with high number of TILs and better prognosis indicates that immunosuppression by M2 macrophages is not necessarily dampening adaptive immune responses but instead kee** them in control to avoid excess inflammation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Common lymphatic endothelial and vascular endothelial receptor-1 (Clever-1, also called Stabilin-1 and Feel-1) is a type I transmembrane protein commonly expressed on non-continuous endothelium [1, 2] and a subset of alternatively activated M2 macrophages [3, 4]. In macrophages, Clever-1 is a scavenger receptor [4, 5] that participates in the scavenging of pathogens, apoptotic cells and molecules [6]. In the endothelium it takes part in lymphocyte, granulocyte and monocyte trafficking and transmigration and can participate in cancer cell migration in the lymphatics [7]. It can also have functions that modulate the immune response [8, 9] and angiogenesis [6]. The gene encoding Clever-1 is called STAB1 [10].
Macrophages are the most abundant benign cells in tumors [11]. Clever-1 is expressed on a subpopulation of alternatively activated macrophages that are immunosuppressive [12]. M2 macrophages are thought to be protumorigenic and essential for tumor progression. They enhance tumor invasiveness, angiogenesis and intravasation into the circulation [11, 13] and stimulate tumor cell proliferation [14]. They are anti-inflammatory and function by negatively affecting cytotoxic CD8 + T-cells [15, 16], and by promoting immunosuppressive regulatory T lymphocytes [17]. They inhibit proinflammatory Th1 type responses and promote Th2 type responses [12]. Clever-1 may participate in many of these functions directly, [12, 13, 15, 16] and therefore anti-Clever therapies are under development [18].
Clever-1 + macrophages have been recorded in solid cancers such as melanoma [19] and glioblastoma [20]. They associate with a worse prognosis in bladder carcinoma [21,22,23] and predict metastasis and risk of recurrence in oral cavity squamous cell carcinoma [24]. Clever-1 + macrophages in gastric cancer predict worse prognosis in tumors with a high T-lymphocyte count [25] or in early stage tumors [26]. High numbers of peritumoral Clever-1 + macrophages in colorectal carcinomas correlate with a better prognosis in early stages, but in the metastatic stage, high intratumoral counts correlate with a worse prognosis [27]. Similarly, one study on bladder cancer has shown that only intratumoral, but not stromal, Clever-1 + macrophages correlate with a worse prognosis [28]. Thus, the evidence on the effect of Clever-1 + macrophages to prognosis is conflicting.
Very little is known about how Clever-1 + macrophages affect breast cancer prognosis and how they associate with different types of breast cancer. Clever-1 + macrophages were reported in 82% of breast carcinomas [29]. The removal of Clever-1 + macrophages inhibit growth of mammary adenocarcinoma in a mouse model [30]. One study showed no correlation between Clever-1 + macrophages and lymph node metastasis [31]. However, another study recorded a correlation between the amount of Clever-1 + macrophages in areas of dense fibrotic stroma in neoadjuvant treated breast cancer and the number of metastatic lymph nodes [32].
We have immunohistochemically investigated 373 cases of primary breast carcinoma for Clever-1 + macrophages. The first aim of this study was to correlate Clever-1 + macrophage numbers with the clinicopathologic features of breast cancer. The second aim was to investigate the relationship of Clever-1 + macrophages with tumor infiltrating lymphocytes (TILs). The third aim was to investigate how Clever-1 + macrophages affect prognosis.
Methods
Patients and tissue samples
The material consisted of all operated primary breast carcinomas in Helsinki University Hospital for the years 1991–1992. All cases were reviewed for tissue sufficiency and after removal of scarce samples, 373 cases remained, for information about the cases see Table 1.
After the selection of cases Hematoxylin and Eosin (H&E)-stained sections were prepared from the most representative blocks and scanned into digital images with Pannoramic Scan 150, Pannoramic Scan II or Pannoramic 250 Flash III (3DHISTECH, Budapest, Hungary). Images were digitally annotated with CaseViewer (3DHISTECH, Budapest, Hungary). Two 1 mm punches were annotated from the center and two from the periphery of each tumor. If the material was scarce, fewer annotations were made. The images were imported and subjected to software analysis, then overlayed with the donor block images, subsequently the layout of the Tissue Microarray (TMA) blocks was designed. TMA Grand Master (3DHISTECH, Budapest, Hungary) tissue microarrayer transferred the cores from the donor block to the recipient TMA blocks, where they were heat-sealed.
Immunohistochemistry
Formalin-fixed paraffin-embedded tissue blocks were cut into 4-µm-thick sections. After deparaffinization, the slides were pretreated in a PT module (LabVision UK Ltd., Suffolk, UK) in Tris–EDTA pH 9.0 (100 °C for 24 min) and cooled to room temperature. Immunohistochemical stainings were done using the following antibodies: CD4 (dilution 1:500, clone 4B12, M7310, Dako), CD8 (dilution 1:100, clone C8/144B, M7103, Dako), ER-alfa (dilution 1:200, clone 6F11, MA5-13,304, Thermo fisher), PR-alfa (dilution 1:200, clone 636, M3569, Dako) and Ki67-1 (dilution 1:75, clone MIB1, M7240 DAKO). The polymer detection kit EnVision (K5007, Dako) was used in a LabVision Autostainer (Thermo Scientific, Fremont, CA). For Clever-1-staining proteinase K antigen retrieval and 2–7 AB (rat IgG) hybridoma medium (in-house, undiluted) was used as previously described [1, 6].
Slides for CD3 and Her2 were stained in Ventana Benchmark Ultra (Ventana/Roche, Tucson, AZ) using the following antibodies: CD3 (RTU, clone 2GV6, 790–4341, Ventana/Roche), HER2 (dilution 1:400, clone CB11, Novocastra). Pretreatment was performed with Ventana Cell Conditioning Solution CC1 (Roche, Tucson, AZ) at 98 °C for 64 min. The primary antibodies were incubated at 36 °C for 32–48 min (Her2 for 48 min, and CD3 for 40 min). OptiView DAB IHC Detection Kit (760–700 Ventana/Roche) was used for detection. The slides were counterstained with Mayer's hematoxylin and mounted in a mounting medium.
All tumors were tested for HER2 gene amplification using Inform HER2 Dual ISH in situ hybridization with Ventana Bechmark Ultra (Ventana/Roche, Tuscon, AZ). After triple pretreatment with solutions CC1 16 at 98 °C for 16 min (950–224, Ventana/Roche) + CC2 at 98 °C for 24 min (950-223, Ventana/Roche) and protease-3 at 37 °C for 16 min (780–4149, Ventana/Roche), the HER2 gene was targeted using a dinitrophenyl labeled probe and the chromosome 17 centromere was localized with a digoxigenin labeled probe (INFORM HER2 Dual ISH DNA Probe Cocktail, 780–4422, Roche/Ventana/Tuscon, AZ, USA 780-4422). HER2 was visualized as black signals with VENTANA ultraView Silver ISH DNP (SISH) Detection (760–098, Roche/Ventana/Tuscon, AZ, USA) and Chr17 as red signals with VENTANA ultraView Red ISH DIG Detection (780–4422, Roche/Ventana/Tuscon, AZ, USA).
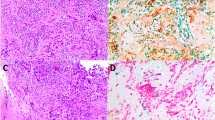
Clever-1 + macrophages were counted using the CaseViewer (3DHISTECH, Budapest, Hungary). Intratumoral and peritumoral Clever-1 + macrophages were counted separately by two persons blinded to each other's counts and from these separate counts mean numbers were calculated (Fig. 1).

Clever-1 stainings. a Punch that was counted to have 37 Clever + positive macrophages. b Punch that was counted to have 490 Clever + positive macrophages
CD3 + , CD4 + and CD8 + cells were evaluated according to and modifying the principles presented by the International TILs working group [33] and the International Immuno-Oncology Biomarkers Working Group [34]. Stromal scores were given by the above- mentioned working groups principles for H&E-sections and intratumoral and peritumoral scores were given by modifying these principles for each compartment. Intratumoral was defined as lymphocytes in direct contact with tumor cells, in contrast to stromal lymphocytes that reside in the tumor stroma without direct contact to tumor cells. Intratumoral (i) CD3 + and CD8 + percentages were defined as low if 0–3%, moderate if 4–9% and high if over 10%. Stromal (s) and peritumoral (p) CD3 + and CD8 + percentages were defined as low if 0–10%, moderate if 11–49% and high if 50–100%. As the CD4 + percentages were generally low, they were categorized into only two categories, low was defined as 0% for iCD4 + and 0–3% for sCD4 + and pCD4 + , high as 1–100% for iCD4 + and 4 –100% for sCD4 + and pCD4 + .
ER and PR were defined as negative when < 1% of tumor cell nuclei were positive and positive when ≥ 1% of tumor cell nuclei were positive. Ki-67 was categorized as being low when ≤ 20% of tumor cell nuclei in hot spots were positive, and high when > 20% were positive. Cases with positivity in HER2 in situ staining were categorized as positive for HER2.
Statistics
Clever-1 + macrophage quantities were correlated with dichotomic clinicopathologic factors and CD4 + cell percentages using the Mann–Whitney-U-Test and with other clinicopathologic factors and CD3 + and CD8 + cell percentages using the Kruskal–Wallis Test. Cumulative survival was calculated using the Kaplan–Meier survivorship method and was analyzed by the log-rank test. A multivariate Cox proportional hazards model was used to estimate the adjusted hazard ratios with 95% confidence intervals (CIs), and to identify independent prognostic factors. P-values < 0.05 were considered significant.
Results
Clever-1 + macrophages according to breast cancer clinicopathologic factors
Means of the numbers of Clever-1 + cells in tumors according to different clinicopathological factors and the p-values for the significance of their differences are shown in Table 2.
ER- tumors had more Clever-1 + macrophages both intra- and peritumorally. There were no significant differences between PR- and PR + tumors or HER2- and HER2 + tumors.
TN tumors had more Clever-1 + macrophages peritumorally than ER + HER- tumors (p = 0.001). No significant differences were seen in pairwise comparisons between the other tumor types, nor regarding the amounts of Clever-1 + macrophages intratumorally.
The Ki-67 high tumors had more Clever-1 + macrophages intra- and peritumorally than the Ki-67 low tumors.
Ductal (p = 0.001) and other types (p = 0.002) of carcinomas had significantly more peritumoral Clever-1 + macrophages than lobular carcinomas, but the difference between ductal and other types of carcinomas was not significant. Grade (G) 3 tumors had significantly more peritumoral Clever-1 + cells than G1 (p < 0.001) and G2 tumors (p = 0.001). There were no differences in the amounts of intratumoral Clever-1 + macrophages.
Tumors > 20 mm had more intra- and peritumoral Clever-1 + macrophages than smaller tumors.
Age of the patient, cancer stage or lymph node status at diagnosis did not correlate with Clever-1 + amounts.
Clever-1 + macrophages correlate with CD3 + , CD8 + and CD4 + lymphocytes
Mean numbers of Clever-1 + macrophages according to CD3 + , CD8 + and CD4 + percentages and p-values for the significance of the differences can be seen in Table 3.
Tumors that had high percentages of iCD3 + cells had higher amounts of intratumoral Clever-1 + macrophages. In the pairwise comparisons, the difference was significant between those tumors with a low percentage of iCD3 + cells and those with a moderate percentage (p = 0.023). Peritumorally the overall differences were significant, but in pairwise comparisons no significant differences were recorded.
Higher iCD8 + percentages also meant higher Clever-1 + cell numbers. This was seen for overall intratumoral Clever-1 + cells, although this difference was not seen in pairwise comparisons. Tumors with moderate amounts of iCD8 + cells had significantly more Clever-1 + cells than tumors with low iCD8 + percentages (p = 0.017).
Tumors that had high percentages of sCD3 + cells also had higher numbers of both intratumoral and peritumoral Clever-1 + macrophages. Moreover, the difference was significant in comparisons with both tumors with a moderate percentage compared to low percentage (p = 0.009 for intratumoral and p = 0.002 for peritumoral) and high percentage compared to low (p = 0.001 for intratumoral and p = 0.001 for peritumoral). The same relationship was seen between sCD8 + cells and Clever-1 + macrophages, this difference was significant in pairwise comparisons only between tumors with a low and moderate percentage of sCD8 + cells (p = 0.003 for intratumoral and p = 0.009 for peritumoral Clever-1 + macrophages).
Higher numbers of Clever-1 + macrophages were seen peritumorally in tumors with higher percentages of pCD3 + and pCD8 + . This difference was significant between tumors with low and moderate percentages of pCD3 + cells (p < 0.001) and pCD8 + cells (p < 0.001) and low and high percentages of pCD3 + cells (p = 0.001). There were no significant differences in the numbers of Clever-1 + macrophages intratumorally.
The Clever-1 + macrophage amounts were higher in tumors with higher amounts of CD4 + cells. This was seen in all compartments for CD4 + cells and for intratumoral and peritumoral Clever-1 + macrophages.
Abundance of intratumoral Clever-1 + macrophages are a favourable prognostic sign
Kaplan–Meier curves that show patient disease-specific survival (DSS) and disease-free survival (DFS) in comparison to intratumoral and peritumoral Clever-1 + amounts are seen in Fig. 2.

Kaplan–Meier curves that show patient disease-specific survival (DSS) and disease-free survival (DFS) in comparison to intratumoral and peritumoral Clever-1 + amounts. Figure created with SPSS 22.0 for Windows. a DSS in months according to intratumoral Clever-1 + amount p = 0.036. b DSS in months according to peritumoral Clever-1 + amount p = 0.749. c DFS in months according to intratumoral Clever-1 + amount p = 0.600. d DFS in months according to peritumoral Clever-1 + amount p = 0.709. e DFS in months according to intratumoral Clever-1 + amount in patients with high percentages of pCD3 p = 0.020. f DFS in months according to intratumoral Clever-1 + amount in patients with high percentages of pCD4 p = 0.008
Patients with low amounts of intratumoral Clever-1 + macrophages had a shorter DSS than patients with higher amounts. No differences were seen in DFS in the entire population, but when analyzed separately patients with high amounts of pCD3 + cells or pCD4 + cells along with low amounts of Clever-1 + cells had shorter DFS. Peritumoral Clever-1 + macrophage counts did not affect survival.
For survival analysis, see Table 4. Univariate Cox proportional hazard analysis was done for Clever-1 + cells, TILs and clinicopathologic factors, and in these analyses intratumoral Clever-1 + cells, sCD8 + cells, sCD4 + cells, stage, size of tumor and lymph node status at diagnosis emerged as prognostic factors for DSS. These factors were chosen for multivariate Cox proportional hazard analysis. In this analysis large tumor size and low amounts of sCD8 + cells and intratumoral Clever-1 + cells were independent adverse prognostic signs.
In a similar manner, univariate analysis was performed for DFS. In this analysis Clever-1 + macrophages were not found to be of prognostic significance. Tumor size, stage, lymph node status and patient age at diagnosis, grade, PR status, sCD8 + cells, pCD8 + cells and sCD4 + cells were prognostic factors and chosen for multivariate analysis. In multivariate analysis, independent factors for shorter DFS were size over 20 mm, younger age, PR negativity and low amounts of pCD8 + cells.
Discussion
We studied Clever-1 + macrophages in 373 breast cancer specimens. To our knowledge this is the first study to investigate Clever-1 + macrophages in a large cohort of breast cancer cases with a long follow-up time. It therefore provides many interesting insights into the distribution and significance of Clever-1 + macrophages and in breast cancer.
We showed that intratumoral Clever-1 + macrophages correlated with a better DSS, whereas peritumoral Clever-1 + macrophages did not affect prognosis. Although Clever-1 + macrophages generally have been associated with a worse prognosis [21,22,23,24, 26], peritumoral Clever-1 + macrophages have been associated with a better prognosis in colorectal carcinoma in early stage [27]. The location of Clever-1 + macrophages seem to be important as intratumoral, unlike stromal, Clever-1 + macrophages have been associated with a higher grade and stage and worse prognosis in bladder cancer [28].
One explanation for why intratumoral Clever-1 + macrophages are associated with a better prognosis, may be related to the function of Clever-1 as a scavenger receptor. Acting locally by scavenging some extracellular components and secreting others, Clever-1 modulates the tumor stroma and its growth milieu.
Clever-1 mediates the endocytosis and lysosomal degradation of Secreted Protein Acidic and Rich in Cysteine (SPARC). SPARC is a soluble extracellular matrix protein that participates in tissue remodeling and binds growth factors thereby inhibiting their function [3, 4, 35]. By affecting the extracellular matrix composition SPARC can potentially affect cancer growth [20]. Research has shown that greater amounts of SPARC in breast cancer in mice inhibit cancer growth [30, 36]. On the other hand, SPARC downregulation in malignant gliomas inhibits cancer cell migration and invasiveness [20] and in lung cancer SPARC has been associated with increased growth and metastatic potential [36]. The exact mechanisms and effect of SPARC is therefore unknown. Clever-1 also participates in the recycling and secretion of chitinase-like proteins that have properties of both cytokines and growth factors. YKL-39 is one of these and is associated with a worse prognosis [37], whereas another, SI-CLP, seems to inhibit growth of mammary adenocarcinoma in mouse models [38]. Yet other target substances of Clever-1 exist and can participate in Clever-modulated ECM remodeling affecting the cancer microenvironment.
On the other hand, Clever-1 is a marker of M2 alternatively activated macrophages. These macrophages are thought to be protumorigenic. Higher amounts of CD68 + cells in breast cancer have been associated with a worse prognosis and M2 macrophages are known to stimulate cancer proliferation, angiogenesis and immunosuppression. The effects of macrophages on prognosis have, however, been contradictory [39]. A lack of Clever-1 has been associated with slower cancer growth in mouse models [17]. This seems contradictory to the association of Clever-1 + macrophages in this study with a better prognosis. It is still worth remembering that only a third of type M2 Tumor Associated Macrophages (TAMs) in cancer are Clever-1 + and Clever is a late marker of M2 macrophage polarization [13, 17].
There are many known effects of Clever-1 + cells and M2 macrophages on the cancer immune reaction. Type − 2 macrophages induce immunosuppressive regulatory T-cells (Tregs) by direct contact and with cytokines [17] and they promote an immunosuppressive Th2 type anti-inflammatory reaction [12]. Anti-Clever-1-therapies have enhanced antitumor CD8 + reactions [15, 18]. According to some studies, Clever-1 directly participates in this process, instead of just being a marker of M2 polarization [12, 15]. Clever-1 + macrophages might bind to CD8 + and CD20 + lymphocytes and thereby inhibit their function. In TNBC with STAB1 overexpression, there seems to be a dysfunction of CD8 + lymphocytes and in some cancers, which are STAB1 high, CD8 + lymphocytes have correlated with a worse prognosis, whereas in cancers with low STAB1, they have correlated with a good prognosis. Additionally Clever-1 + mediated lysosomal degradation of products affects antigen presentation [16]. It is also possible that Clever-1 + macrophages mediate lymphocyte extravasation and accumulation, which enhances tumor cell killing. In this study, the amounts of TILs positively correlated with Clever-1 + macrophages. As intratumoral Clever-1 + macrophages correlated with a better prognosis, it would seem, that they were not able to entirely inhibit the function of cytotoxic T-cells.
CD8 + lymphocytes are cytotoxic cells that engage in tumor killing by inducing cytolysis [40], which explains their positive effect on prognosis, a finding well in line with previous studies [41,42,43,44,45]. PR negativity associates with a worse prognosis in ER + breast cancer, and with resistance to endocrine treatment [46, 47].
In this study, higher amounts of Clever-1 + macrophages correlated with a higher grade, ER negativity and larger size, but not with HER2 status. This finding is well in line with previous studies concerning TAMs in breast cancer [39]. The same tumors also had higher amounts of TILs, and in studies on TILs, they have been associated with many of the same factors as TAMs, although HER2 + tumors tend to have higher amounts of TILs [48, 49]. TILs are associated with a better prognosis in at least TNBC and HER2 + breast cancer [50]. According to our results the same tumors that attract TILs also attract Clever-1 + macrophages, creating an immune-rich phenotype. Clever-1 + macrophages can attract TILs, but they can also modulate their function.
This study has all the limitations known for a retrospective study. TAMs tend to cluster, which limits the accuracy of TMAs in this context [21]. This limitation has been considered, and therefore several sample punches per tumor were taken for study.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
References
Irjala H, Alanen K, Grénman R, Heikkilä P, Joensuu H, Jalkanen S (2003) Mannose receptor (MR) and common lymphatic endothelial and vascular endothelial receptor (CLEVER)-1 direct the binding of cancer cells to the lymph vessel endothelium. Cancer Res 63:4671–4676
Irjala H, Elima K, Johansson EL, Merinen M, Kontula K, Alanen K, Grenman R, Salmi M, Jalkanen S (2003) The same endothelial receptor controls lymphocyte traffic both in vascular and lymphatic vessels. Eur J Immunol 33:815–824. https://doi.org/10.1002/eji.200323859
Kzhyshkowska J (2010) Multifunctional receptor stabilin-1 in homeostasis and disease. ScientificWorldJournal 10:2039–2053. https://doi.org/10.1100/tsw.2010.189
Kzhyshkowska J, Gratchev A, Goerdt S (2006) Stabilin-1, a homeostatic scavenger receptor with multiple functions. J Cell Mol Med 10:635–649. https://doi.org/10.1111/j.1582-4934.2006.tb00425.x
Prevo R, Banerji S, Ni J, Jackson DG (2004) Rapid plasma membrane-endosomal trafficking of the lymph node sinus and high endothelial venule scavenger receptor/homing receptor stabilin-1 (FEEL-1/CLEVER-1). J Biol Chem 279:52580–52592. https://doi.org/10.1074/jbc.M406897200
Palani S, Maksimow M, Miiluniemi M, Auvinen K, Jalkanen S, Salmi M (2011) Stabilin-1/CLEVER-1, a type 2 macrophage marker, is an adhesion and scavenging molecule on human placental macrophages. Eur J Immunol 41:2052–2063. https://doi.org/10.1002/eji.201041376
Salmi M, Koskinen K, Henttinen T, Elima K, Jalkanen S (2004) CLEVER-1 mediates lymphocyte transmigration through vascular and lymphatic endothelium. Blood 104:3849–3857. https://doi.org/10.1182/blood-2004-01-0222
Shetty S, Weston CJ, Oo YH, Westerlund N, Stamataki Z, Youster J, Hubscher SG, Salmi M, Jalkanen S, Lalor PF, Adams DH (2011) Common lymphatic endothelial and vascular endothelial receptor-1 mediates the transmigration of regulatory T cells across human hepatic sinusoidal endothelium. J Immunol 186:4147–4155. https://doi.org/10.4049/jimmunol.1002961
Tadayon S, Dunkel J, Takeda A, Eichin D, Virtakoivu R, Elima K, Jalkanen S, Hollmén M (2021) Lymphatic endothelial cell activation and dendritic cell transmigration is modified by genetic deletion of clever-1. Front Immunol 12:602122. https://doi.org/10.3389/fimmu.2021.602122
Dunkel J, Viitala M, Karikoski M, Rantakari P, Virtakoivu R, Elima K, Hollmén M, Jalkanen S, Salmi M (2018) Enhanced antibody production in clever-1/stabilin-1-deficient mice. Front Immunol 9:2257. https://doi.org/10.3389/fimmu.2018.02257
Noy R, Pollard JW (2014) Tumor-associated macrophages: from mechanisms to therapy. Immunity 41:49–61. https://doi.org/10.1016/j.immuni.2014.06.010
Palani S, Elima K, Ekholm E, Jalkanen S, Salmi M (2016) Monocyte stabilin-1 suppresses the activation of Th1 lymphocytes. J Immunol 196:115–123. https://doi.org/10.4049/jimmunol.1500257
Riabov V, Gudima A, Wang N, Mickley A, Orekhov A, Kzhyshkowska J (2014) Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Front Physiol 5:75. https://doi.org/10.3389/fphys.2014.00075
Rasmussen RK, Etzerodt A (2021) Therapeutic targeting of tumor-associated macrophages. Adv Pharmacol 91:185–211. https://doi.org/10.1016/bs.apha.2021.03.002
Viitala M, Virtakoivu R, Tadayon S, Rannikko J, Jalkanen S, Hollmén M (2019) Immunotherapeutic blockade of macrophage clever-1 reactivates the CD8(+) T-cell response against immunosuppressive tumors. Clin Cancer Res 25:3289–3303. https://doi.org/10.1158/1078-0432.Ccr-18-3016
Hollmén M, Figueiredo CR, Jalkanen S (2020) New tools to prevent cancer growth and spread: a “Clever” approach. Br J Cancer 123:501–509. https://doi.org/10.1038/s41416-020-0953-0
Karikoski M, Marttila-Ichihara F, Elima K, Rantakari P, Hollmén M, Kelkka T, Gerke H, Huovinen V, Irjala H, Holmdahl R, Salmi M, Jalkanen S (2014) Clever-1/stabilin-1 controls cancer growth and metastasis. Clin Cancer Res 20:6452–6464. https://doi.org/10.1158/1078-0432.Ccr-14-1236
Virtakoivu R, Rannikko JH, Viitala M, Vaura F, Takeda A, Lönnberg T, Koivunen J, Jaakkola P, Pasanen A, Shetty S, de Jonge MJA, Robbrecht D, Ma YT, Skyttä T, Minchom A, Jalkanen S, Karvonen MK, Mandelin J, Bono P, Hollmén M (2021) Systemic blockade of clever-1 elicits lymphocyte activation alongside checkpoint molecule downregulation in patients with solid tumors: results from a phase I/II clinical trial. Clin Cancer Res 27:4205–4220. https://doi.org/10.1158/1078-0432.Ccr-20-4862
Schönhaar K, Schledzewski K, Michel J, Dollt C, Gkaniatsou C, Géraud C, Kzhyshkowska J, Goerdt S, Schmieder A (2014) Expression of stabilin-1 in M2 macrophages in human granulomatous disease and melanocytic lesions. Int J Clin Exp Pathol 7:1625–1634
David C, Nance JP, Hubbard J, Hsu M, Binder D, Wilson EH (2012) Stabilin-1 expression in tumor associated macrophages. Brain Res 1481:71–78. https://doi.org/10.1016/j.brainres.2012.08.048
Tervahartiala M, Taimen P, Mirtti T, Koskinen I, Ecke T, Jalkanen S, Boström PJ (2017) Immunological tumor status may predict response to neoadjuvant chemotherapy and outcome after radical cystectomy in bladder cancer. Sci Rep 7:12682. https://doi.org/10.1038/s41598-017-12892-5
Zhang Q, Mao Z, Sun J (2019) NF-κB inhibitor, BAY11-7082, suppresses M2 tumor-associated macrophage induced EMT potential via miR-30a/NF-κB/Snail signaling in bladder cancer cells. Gene 710:91–97. https://doi.org/10.1016/j.gene.2019.04.039
Ecke TH, Kiani A, Schlomm T, Friedersdorff F, Rabien A, Jung K, Kilic E, Boström P, Tervahartiala M, Taimen P, Gleichenhagen J, Johnen G, Brüning T, Koch S, Roggisch J, Wirtz RM (2020) Prognostic role of survivin and macrophage infiltration quantified on protein and mRNA level in molecular subtypes determined by RT-qPCR of KRT5, KRT20, and ERBB2 in muscle-invasive bladder cancer treated by adjuvant chemotherapy. Int J Mol Sci. https://doi.org/10.3390/ijms21197420
Kwon M, Yeo SC, Lee JS, Park JJ (2019) Not CD68 but stabilin-1 expression is associated with the risk of recurrence in patients with oral cavity squamous cell carcinoma. Head Neck 41:2058–2064. https://doi.org/10.1002/hed.25654
Junttila A, Helminen O, Väyrynen JP, Ahtiainen M, Kenessey I, Jalkanen S, Mecklin JP, Kellokumpu I, Kuopio T, Böhm J, Mrena J (2020) Immunophenotype based on inflammatory cells, PD-1/PD-L1 signalling pathway and M2 macrophages predicts survival in gastric cancer. Br J Cancer 123:1625–1632. https://doi.org/10.1038/s41416-020-01053-7
Yin SP, Gao Y, **e XS, Xu DD, Riabov V, Du WD (2020) Accumulation of stabilin-1 positive macrophages in the early stage of gastric cancer is associated with short cumulative survival. Oncol Lett 19:2404–2412. https://doi.org/10.3892/ol.2020.11310
Algars A, Irjala H, Vaittinen S, Huhtinen H, Sundström J, Salmi M, Ristamäki R, Jalkanen S (2012) Type and location of tumor-infiltrating macrophages and lymphatic vessels predict survival of colorectal cancer patients. Int J Cancer 131:864–873. https://doi.org/10.1002/ijc.26457
Wang B, Huang H, Yang M, Yang W, Liu Z, Hou W, Zeng H, He Z, Lin T, Huang J (2020) Microlocalization and clinical significance of stabilin-1(+) macrophages in treatment-naïve patients with urothelial carcinoma of the bladder. World J Urol 38:709–716. https://doi.org/10.1007/s00345-019-02853-0
Ammar A, Mohammed RA, Salmi M, Pepper M, Paish EC, Ellis IO, Martin SG (2011) Lymphatic expression of CLEVER-1 in breast cancer and its relationship with lymph node metastasis. Anal Cell Pathol (Amst) 34:67–78. https://doi.org/10.3233/acp-2011-0002
Riabov V, Yin S, Song B, Avdic A, Schledzewski K, Ovsiy I, Gratchev A, Llopis Verdiell M, Sticht C, Schmuttermaier C, Schönhaber H, Weiss C, Fields AP, Simon-Keller K, Pfister F, Berlit S, Marx A, Arnold B, Goerdt S, Kzhyshkowska J (2016) Stabilin-1 is expressed in human breast cancer and supports tumor growth in mammary adenocarcinoma mouse model. Oncotarget 7:31097–31110. https://doi.org/10.18632/oncotarget.8857
Buldakov M, Zavyalova M, Krakhmal N, Telegina N, Vtorushin S, Mitrofanova I, Riabov V, Yin S, Song B, Cherdyntseva N, Kzhyshkowska J (2017) CD68+, but not stabilin-1+ tumor associated macrophages in gaps of ductal tumor structures negatively correlate with the lymphatic metastasis in human breast cancer. Immunobiology 222:31–38. https://doi.org/10.1016/j.imbio.2015.09.011
Mitrofanova I, Zavyalova M, Telegina N, Buldakov M, Riabov V, Cherdyntseva N, Kzhyshkowska J (2017) Tumor-associated macrophages in human breast cancer parenchyma negatively correlate with lymphatic metastasis after neoadjuvant chemotherapy. Immunobiology 222:101–109. https://doi.org/10.1016/j.imbio.2016.08.001
Salgado R, Denkert C, Demaria S, Sirtaine N, Klauschen F, Pruneri G, Wienert S, Van den Eynden G, Baehner FL, Penault-Llorca F, Perez EA, Thompson EA, Symmans WF, Richardson AL, Brock J, Criscitiello C, Bailey H, Ignatiadis M, Floris G, Sparano J, Kos Z, Nielsen T, Rimm DL, Allison KH, Reis-Filho JS, Loibl S, Sotiriou C, Viale G, Badve S, Adams S, Willard-Gallo K, Loi S (2015) The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: recommendations by an International TILs Working Group 2014. Ann Oncol 26:259–271. https://doi.org/10.1093/annonc/mdu450
Hendry S, Salgado R, Gevaert T, Russell PA, John T, Thapa B, Christie M, van de Vijver K, Estrada MV, Gonzalez-Ericsson PI, Sanders M, Solomon B, Solinas C, Van den Eynden GGGM, Allory Y, Preusser M, Hainfellner J, Pruneri G, Vingiani A, Demaria S, Symmans F, Nuciforo P, Comerma L, Thompson EA, Lakhani S, Kim S-R, Schnitt S, Colpaert C, Sotiriou C, Scherer SJ, Ignatiadis M, Badve S, Pierce RH, Viale G, Sirtaine N, Penault-Llorca F, Sugie T, Fineberg S, Paik S, Srinivasan A, Richardson A, Wang Y, Chmielik E, Brock J, Johnson DB, Balko J, Wienert S, Bossuyt V, Michiels S, Ternes N, Burchardi N, Luen SJ, Savas P, Klauschen F, Watson PH, Nelson BH, Criscitiello C, O’Toole S, Larsimont D, de Wind R, Curigliano G, André F, Lacroix-Triki M, van de Vijver M, Rojo F, Floris G, Bedri S, Sparano J, Rimm D, Nielsen T, Kos Z, Hewitt S, Singh B, Farshid G, Loibl S, Allison KH, Tung N, Adams S, Willard-Gallo K, Horlings HM, Gandhi L, Moreira A, Hirsch F, Dieci MV, Urbanowicz M, Brcic I, Korski K, Gaire F, Koeppen H, Lo A, Giltnane J, Rebelatto MC, Steele KE, Zha J, Emancipator K, Juco JW, Denkert C, Reis-Filho J, Loi S, Fox SB (2017) Assessing tumor-infiltrating lymphocytes in solid tumors: a practical review for pathologists and proposal for a standardized method from the international immunooncology biomarkers working group: part 1: assessing the host immune response, TILs in invasive breast carcinoma and ductal carcinoma in situ, metastatic tumor deposits and areas for further research. Adv Anat Pathol 24:235–251. https://doi.org/10.1097/PAP.0000000000000162
Karikoski M, Irjala H, Maksimow M, Miiluniemi M, Granfors K, Hernesniemi S, Elima K, Moldenhauer G, Schledzewski K, Kzhyshkowska J, Goerdt S, Salmi M, Jalkanen S (2009) Clever-1/stabilin-1 regulates lymphocyte migration within lymphatics and leukocyte entrance to sites of inflammation. Eur J Immunol 39:3477–3487. https://doi.org/10.1002/eji.200939896
Kzhyshkowska J, Workman G, Cardó-Vila M, Arap W, Pasqualini R, Gratchev A, Krusell L, Goerdt S, Sage EH (2006) Novel function of alternatively activated macrophages: stabilin-1-mediated clearance of SPARC. J Immunol 176:5825–5832. https://doi.org/10.4049/jimmunol.176.10.5825
Liu T, Larionova I, Litviakov N, Riabov V, Zavyalova M, Tsyganov M, Buldakov M, Song B, Moganti K, Kazantseva P, Slonimskaya E, Kremmer E, Flatley A, Klüter H, Cherdyntseva N, Kzhyshkowska J (2018) Tumor-associated macrophages in human breast cancer produce new monocyte attracting and pro-angiogenic factor YKL-39 indicative for increased metastasis after neoadjuvant chemotherapy. Oncoimmunology 7:e1436922. https://doi.org/10.1080/2162402x.2018.1436922
Yin S, Wang N, Riabov V, Mossel DM, Larionova I, Schledzewski K, Trofimova O, Sevastyanova T, Zajakina A, Schmuttermaier C, Gratchev A, Flatley A, Kremmer E, Zavyalova M, Cherdyntseva N, Simon-Keller K, Marx A, Klüter H, Goerdt S, Kzhyshkowska J (2020) SI-CLP inhibits the growth of mouse mammary adenocarcinoma by preventing recruitment of tumor-associated macrophages. Int J Cancer 146:1396–1408. https://doi.org/10.1002/ijc.32685
Larionova I, Tuguzbaeva G, Ponomaryova A, Stakheyeva M, Cherdyntseva N, Pavlov V, Choinzonov E, Kzhyshkowska J (2020) Tumor-associated macrophages in human breast, colorectal, lung. Ovarian Prostate Cancers Front Oncol 10:566511. https://doi.org/10.3389/fonc.2020.566511
Pruneri G, Vingiani A, Denkert C (2018) Tumor infiltrating lymphocytes in early breast cancer. Breast (Edinburgh, Scotland) 37:207–214. https://doi.org/10.1016/j.breast.2017.03.010
Vihervuori H, Autere TA, Repo H, Kurki S, Kallio L, Lintunen MM, Talvinen K, Kronqvist P (2019) Tumor-infiltrating lymphocytes and CD8+ T cells predict survival of triple-negative breast cancer. J Cancer Res Clin Oncol 145:3105–3114. https://doi.org/10.1007/s00432-019-03036-5
Egelston CA, Avalos C, Tu TY, Rosario A, Wang R, Solomon S, Srinivasan G, Nelson MS, Huang Y, Lim MH, Simons DL, He T-F, Yim JH, Kruper L, Mortimer J, Yost S, Guo W, Ruel C, Frankel PH, Yuan Y, Lee PP (2019) Resident memory CD8+ T cells within cancer islands mediate survival in breast cancer patients. JCI insight. https://doi.org/10.1172/jci.insight.130000
Ali HR, Provenzano E, Dawson SJ, Blows FM, Liu B, Shah M, Earl HM, Poole CJ, Hiller L, Dunn JA, Bowden SJ, Twelves C, Bartlett JMS, Mahmoud SMA, Rakha E, Ellis IO, Liu S, Gao D, Nielsen TO, Pharoah PDP, Caldas C (2014) Association between CD8+ T-cell infiltration and breast cancer survival in 12,439 patients. Ann Oncol 25:1536–1543. https://doi.org/10.1093/annonc/mdu191
Liu S, Lachapelle J, Leung S, Gao D, Foulkes WD, Nielsen TO (2012) CD8+ lymphocyte infiltration is an independent favorable prognostic indicator in basal-like breast cancer. Breast Cancer Res 14:R48. https://doi.org/10.1186/bcr3148
Mahmoud SMA, Paish EC, Powe DG, Macmillan RD, Grainge MJ, Lee AHS, Ellis IO, Green AR (2011) Tumor-infiltrating CD8+ lymphocytes predict clinical outcome in breast cancer. J Clin Oncol 29:1949–1955. https://doi.org/10.1200/JCO.2010.30.5037
Li Z, Wei H, Li S, Wu P, Mao X (2022) The role of progesterone receptors in breast cancer. Drug Des Devel Ther 16:305–314. https://doi.org/10.2147/dddt.S336643
Cuyún Carter G, Mohanty M, Stenger K, Morato Guimaraes C, Singuru S, Basa P, Singh S, Tongbram V, Kuemmel S, Guarneri V, Tolaney SM (2021) Prognostic factors in hormone receptor-positive/human epidermal growth factor receptor 2-negative (HR+/HER2-) advanced breast cancer: a systematic literature review. Cancer Manag Res 13:6537–6566. https://doi.org/10.2147/cmar.S300869
Solinas C, Carbognin L, De Silva P, Criscitiello C, Lambertini M (2017) Tumor-infiltrating lymphocytes in breast cancer according to tumor subtype: current state of the art. Breast (Edinburgh, Scotland) 35:142–150. https://doi.org/10.1016/j.breast.2017.07.005
Denkert C, von Minckwitz G, Darb-Esfahani S, Lederer B, Heppner BI, Weber KE, Budczies J, Huober J, Klauschen F, Furlanetto J, Schmitt WD, Blohmer J-U, Karn T, Pfitzner BM, Kümmel S, Engels K, Schneeweiss A, Hartmann A, Noske A, Fasching PA, Jackisch C, van Mackelenbergh M, Sinn P, Schem C, Hanusch C, Untch M, Loibl S (2018) Tumour-infiltrating lymphocytes and prognosis in different subtypes of breast cancer: a pooled analysis of 3771 patients treated with neoadjuvant therapy. Lancet Oncol 19:40–50. https://doi.org/10.1016/S1470-2045(17)30904-X
Mao Y, Qu Q, Chen X, Huang O, Wu J, Shen K (2016) The prognostic value of tumor-infiltrating lymphocytes in breast cancer: a systematic review and meta-analysis. PLoS ONE 11:e0152500. https://doi.org/10.1371/journal.pone.0152500
Acknowledgements
We warmly thank Eija Heiliö and Sari Mäki for excellent technical assistance. We thank Mia Kero for sharing her expertise on immunohistochemistry. We thank Alisdair Mclean for checking the language in this article.
Funding
Open Access funding provided by University of Helsinki including Helsinki University Central Hospital. This study was funded by the Helsinki University Central Hospital Research Foundation and the Academy of Finland (MH).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Maija Hollmen own shares of Faron Pharmaceuticals and reports receiving funding from Faron for the preclinical development of anti-Clever-1 mAbs. The authors have no other relevant financial or non-financial interests to disclose.
Ethical approval
This study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the Ethics Committee of the Helsinki University Central Hospital.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mutka, M., Virtakoivu, R., Joensuu, K. et al. Clever-1 positive macrophages in breast cancer. Breast Cancer Res Treat 195, 237–248 (2022). https://doi.org/10.1007/s10549-022-06683-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10549-022-06683-4