
Abstract
Background
Spinal central sensitization is an important process in the generation and maintenance of visceral hypersensitivity. The release of brain-derived neurotrophic factor (BDNF) from the primary afferent neurons to the spinal cord contributes to spinal neuronal plasticity and increases neuronal activity and synaptic efficacy. The N-Methyl-D-aspartic acid (NMDA) receptor possesses ion channel properties, and its activity is modulated by phosphorylation of its subunits including the NMDA receptor 1 (NR1).
Methods
Colonic inflammation was induced by a single dose of intracolonic instillation of tri-nitrobenzene sulfonic acid (TNBS). NR1 phosphorylation by BDNF in vivo and in culture was examined by western blot and immunohistochemistry. Signal transduction was studied by direct examination and use of specific inhibitors.
Results
During colitis, the level of NR1 phospho-Ser896 was increased in the dorsal horn region of the L1 and S1 spinal cord; this increase was attenuated by injection of BDNF neutralizing antibody to colitic animals (36 μg/kg, intravenous (i.v.)) and was also reduced in BDNF+/− rat treated with TNBS. Signal transduction examination showed that the extracellular signal-regulated kinase (ERK) activation was not involved in BDNF-induced NR1 phosphorylation. In contrast, the phosphatidylinositol 3-kinase (PI3K)/Akt pathway mediated BDNF-induced NR1 phosphorylation in vivo and in culture; this is an additional pathway to the phospholipase C-gamma (PLCγ) and the protein kinase C (PKC) that was widely considered to phosphorylate NR1 at Ser896. In spinal cord culture, the inhibitors to PLC (U73122), PKC (bisindolylmaleimide I), and PI3K (LY294002), but not MEK (PD98059) blocked BDNF-induced NR1 phosphorylation. In animals with colitis, treatment with LY294002 (50 μg/kg, i.v.) blocked the Akt activity as well as NR1 phosphorylation at Ser896 in the spinal cord.
Conclusion
BDNF participates in colitis-induced spinal central sensitization by up-regulating NR1 phosphorylation at Ser896. The PI3K/Akt pathway, in addition to PLCγ and PKC, mediates BDNF action in the spinal cord during colitis.
Similar content being viewed by others

Background
Spinal central sensitization is suggested to be one of the key mechanisms underlying visceral hypersensitivity in response to visceral irritation and/or inflammation in patients and experimental animal models [1–3]. Alterations in the expression and activity levels of neurochemical compounds and ion channels in the spinal dorsal horn underlie the molecular mechanisms of spinal central sensitization and is modulated by release of neurotransmitters from the primary sensory neurons located in the dorsal root ganglia (DRG) [4, 5]. Following colonic inflammation such as colitis induced by chemicals including tri-nitrobenzene sulfonic acid (TNBS), zymosan, acetic acid, mustard oil, or dextran sulfate sodium, there are increased levels of neurotrophins and neuropeptides in the DRG and the spinal cord resulting in visceral hypersensitivity [6–10]. As a result of zymosan-induced colitis, visceral hyperalgesia in rats is mediated, at least in part, by spinal N-methyl-D-aspartate (NMDA) receptor and is attenuated by intrathecal administration of the non-competitive NMDA receptor channel pharmacological blocker [7]. Peripheral tissue and nerve injuries recruit NMDA receptor at synapses within the pain-related spinal dorsal horn which is highly considered as an important process in spinal central sensitization under several physiologic and pathophysiologic conditions [11–16].
The NMDA receptor has an ionotropic property which regulates Ca2+ influx and Ca2+-dependent physiological effects thereby regulating neuronal activity and synaptic efficacy. NMDA receptor forms a heterotetramer composed of two NMDA receptor 1 (NR1) and two NR2 subunits [17]. NMDA receptor function may depend on phosphorylation of the subunits of the NMDA receptor [18, 19]. The NR1 subunit can be phosphorylated at Ser890 and Ser896 by protein kinase C (PKC) and at Ser897 by protein kinase A (PKA) [20, 21]. This is also true in animal models. Phosphorylation of Ser897 of the NR1 subunit in the spinal dorsal horn and spinothalamic tract neurons after intradermal injection of capsaicin in rats is mediated by the PKA pathway and is sensitive to PKA inhibitors [21]. Phosphorylation of Ser896 of the NR1 subunit in the spinal dorsal horn following noxious heat stimulation of the rat hind paw is mediated by activation of PKC [22]. In cystitis, both Ser896 and Ser897 sites of the NR1 subunit are phosphorylated in the spinal cord [5]. The Ser897 but not Ser896 phosphorylation in the spinal cord is regulated by calcitonin gene-related peptide (CGRP) [5]. We speculate that brain-derived neurotrophic factor (BDNF) may be involved in PKC-mediated NR1 Ser896 phosphorylation in the spinal cord during visceral inflammation.
BDNF is a member of nerve growth factor family. The release of BDNF from the neuron somata in the DRG to the spinal dorsal horn, where BDNF binds high affinity receptor TrkB, influences associative synaptic plasticity and increases synaptic efficacy by refinement of neural connectivity [9, 23, 24]. The involvement of BDNF in sensory hypersensitive is proved by a line of evidence and is well discussed in several animal models. In particular with colitis, systemic injection of BDNF neutralizing antibody reverses colonic hypersensitivity in response to colonic distention [25]. Intrathecal infusion of BDNF neutralizing antibody via mini pump also attenuates colitis-associated bladder hyperactivity [10]. In other animal models, the role of BDNF in mediating sensory sensitization is observed during peripheral inflammation-induced somatic pain [26, 27], cancer-induced bone pain [28], nerve injury, etc. [29–32]. Traditionally, three signaling pathways are activated by BDNF in neurons: activation of the Ras-extracellular signal-regulated kinase (ERK) cascade leads to gene transcription promoting cellular growth, the phosphatidylinositol 3-kinase (PI3K)/Akt pathway has anti-apoptotic function, and activation of phospholipase C-gamma (PLCγ) initiates Ca2+ release or influx via modulation of store activity or ion channels possibly through diacyl glycerol (DAG)-mediated PKC activation [33]. These pathways are recently recognized as essential components in sensory hypersensitivity in several animal models [34–38].
In culture, BDNF is able to modulate NMDA receptor in activity-dependent manner [39]. In the spinal cord, BDNF regulates the activity of the NR2B-containing NMDA receptor thereby participating in the development of neuropathic pain [40, 41]. The interrelationship of BDNF with the NR1 subunit of the NMDA receptor has also been reported showing that BDNF can regulate NR1 protein synthesis in vivo and in culture [28, 42]. Earlier studies demonstrate a role of BDNF in inducing NR1 phosphorylation on Ser897, the traditional PKA site, via the ERK and PKC pathways in isolated spinal dorsal horn [43, 44]. The present study is undertaken to examine the possibility of BDNF in regulating the NR1 subunit of the NMDA receptor by phosphorylating Ser896, the traditional PKC site, in vivo and in spinal culture, and explore the signal transduction in an animal model of colitis-induced visceral hypersensitivity.
Experimental procedures
Animals and reagents
Adult male rats (150–200 g) were purchased from Harlan Sprague Dawley, Inc. (Indianapolis, IN). All experimental protocols involving animal use were approved by the Institutional Animal Care and Use Committee at the Virginia Commonwealth University. Animal care was in accordance with the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) and National Institutes of Health guidelines. All efforts were made to minimize the potential for animal pain, stress, or distress as well as to reduce the number of animals used. TNBS, β-actin antibody, and other chemicals used in this experiment were purchased from Sigma-Aldrich (St. Louis, MO). Antibody against p-NR1 Ser896 was from Santa Cruz Biotechnology (Santa Cruz, CA). Antibodies for p-Akt and total Akt were from Cell Signaling Technology (Danvers, MA). Secondary antibodies for western blot were from Pierce Biotechnology (Rockford, IL), and secondary antibodies for immunohistochemistry were from Molecular Probes (Eugene, OR).
Induction of colonic inflammation
Colonic inflammation was induced in fasted rats under anesthesia (2 % isoflurane). TNBS solution (60 mg/mL in 50 % EtOH) at a dose of 90 mg/kg (1.5 mL TNBS solution per kg body weight) was instilled into the lumen of the distal colon through a syringe-attached polyethylene catheter via the rectum 6 cm proximal to the anus. Control animals received similar volume of 50 % EtOH enema. Euthanasia of animals was performed on day 3 or day 7 after TNBS treatment. To ensure exposure of the distal colon to TNBS, rats were held head down by lifting up the tail for 1 min. This treatment regimen caused significant inflammatory responses identified by histology [45] as well as the up-regulation of pro-inflammatory factor in the distal colon (Additional file 1: Supplemental data).
Treatment of animals with antagonists
Animals received antagonists via intravenous (i.v.) injection. Antagonists used in this study were BDNF neutralizing antibody (36 μg/kg body weight) and the PI3K inhibitor LY 294002 (50 μg/kg body weight). A single dose of antagonist was injected. When animals were examined on day 3 post colitis induction, the antagonist was injected on the same day and post-TNBS treatment. When animals were examined on day 7 post colitis induction, the antagonist was injected on day 3 after colitis induction. This treatment design was customized by us through preliminary studies.
Tissue collection
The L1 and S1 spinal cord segments were dissected out and used for western blot, immunostaining, or acute culture. For western blot, spinal segments were freshly minced and homogenized in T-PER buffer (Pierce Biotechnology, Rockford, IL) supplemented with protease and phosphatase inhibitors (Sigma). For immunohistochemistry, spinal segments were fixed with 4 % paraformaldehyde in 0.1 M PBS (pH = 7.4) followed by 20 % sucrose for cryoprotection. For acute culture, spinal segments were freshly dissected out from naïve animals, transversely sectioned at a thickness of 250 μm, and randomly divided into groups for treatment.
Western blot
The protein extracts were subject to centrifugation at 20,200g for 10 min at 4 °C, and the supernatant was removed to a fresh tube. The protein concentration was determined using Bio-Rad DC protein assay kit. Proteins were then separated on a 10 % SDS-PAGE gel and transferred to a nitrocellulose membrane. The membrane was blocked with 5 % milk in Tris-buffered saline for 1 h and then incubated with a specific primary antibody followed by HRP-conjugated secondary antibody. For internal loading control, the same membrane was stripped and re-probed with anti-β-actin antiserum or antibody to a non-phosphorylated form of the protein examined. The concentrations of antibodies used were p-NR1: 1:1000; p-Akt and Akt: 1:1000; and β-actin: 1:3000. The bands were visualized by enhanced chemiluminescence (ECL). Densitometric quantification of the immunoreactive bands was performed using the software FluorChem 8800 (Alpha Innotech, San Leandro, CA).
Immunohistochemistry
The spinal cord segments were sectioned transversely at a thickness of 30 μm and were immunostained by free-floating method. Generally, sections were incubated with blocking solution containing 5 % normal donkey serum (Jackson Immuno Research, West Grove, PA) in PBST (0.3 % Triton X-100 in 0.1 M PBS, pH 7.4) for 30 min followed by specific primary antibodies overnight at 4 °C. After rinsing (3 × 10 min with 0.1 M PBS), tissues were incubated with fluorescence-conjugated species-specific secondary antibody for 2 h at room temperature. Following washing, the sections were mounted to slides and coverslipped with Citifluor (Citifluor Ltd., London). The sections were then viewed and analyzed with a Zeiss AxioImage Z1 Apitome fluorescent microscope.
The analysis of the immunoreactivity at the dorsal horn were done as previously reported by converting fluorescent images to a grayscale that ranged in intensity from 0 (black) to 255 (white) for the purpose of densitometry [46]. The same number of standard sized rectangles was overlaid on the area of interest (i.e., superficial dorsal horn in this study) for each spinal section. Intensity measured within the rectangles was averaged as one point.
Spinal cord culture
Spinal cord segments were acutely cultured for 4–6 h in cell culture wells containing Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 200 units/mL penicillin, 200 mg/mL streptomycin, and 100 mg/mL gentamycin. BDNF (50 ng/mL) was added to the culture medium and incubated for designated time points. After incubation, tissues were collected for western blot analysis. All cultures were maintained in a 10 % CO2 environment at 37 °C.
Statistical analysis
Comparison between control and experimental groups was made by using Kruskal-Wallis non-parametric one-way ANOVA. For in vivo experiments, 4–6 animals were used for each experimental group. For culture, 3–4 independent experiments were performed. Results were presented as mean ± SE. Differences between means at a level of p ≤ 0.05 were considered to be significant.
Results
Up-regulation of NR1 phosphorylation at Ser896 in spinal dorsal horn during colitis
The expression level and the distribution pattern of the phospho (p)-NR1 in the spinal cord were examined by western blot and immunohistochemistry. We used specific antibody that recognized phospho-Ser896 on the NR1 subunit as described previously [5]. Western blot results showed that the level of p-NR1 Ser896 was increased in both L1 (Fig. 1a, b) and S1 (Fig. 1c, d) spinal cord at 3 days and 7 days post colitis induction. Immunohistochemistry staining showed that p-NR1 Ser896 immunoreactivity was expressed in several regions of the spinal cord. We paid particular attention to the dorsal horn region where the primary sensory neuron terminals ended. As shown in Fig. 2, colitis increased the immunodensity of p-NR1 Ser896 in the dorsal horn of the L1 (Fig. 2a, b) and S1 spinal cord (Fig. 2c, d). Summary data (Fig. 2e, f) presented the changes in p-NR1 Ser896 immunoreactivity in the L1 and S1 spinal dorsal horn at both 3 days and 7 days postcolitis. It is noteworthy that we have identified that phosphorylation of Ser897 of the NR1 subunit in the spinal cord was regulated by CGRP [5]. However, CGRP failed to regulate the phosphorylation of Ser896 on the NR1 subunit [5]. Thus, the present study focused on the examination of NR1 phosphorylation at Ser896 and aimed to identify factors that mediated NR1 Ser896 phosphorylation in the spinal cord in an animal model of colitis.

Up-regulation of NR1 phosphorylation in the L1 and S1 spinal cord during colitis. At 3 and 7 days post-TNBS treatment, the level of p-NR1 was examined by western blot (a, c) in the L1 (a, b) and S1 (c, d) spinal cord using a specific antibody against NR1 Ser896. Results showed that colitis caused significantly increases in the level of p-NR1 in the L1 and S1 spinal cord at both 3 days and 7 days post-TNBS treatment. *p < 0.05 vs. control

NR1 phosphorylation was increased in the dorsal horn of the spinal cord during colitis. Up-regulation of p-NR1 in the spinal cord during colitis was examined by immunohistochemistry (a–d). The p-NR1 immunoreactivity was increased in the region of the spinal dorsal horn in L1 (a–b) and S1 (c–d) spinal cord during colitis (compare b to a and d to c, circled area). Histograms (e, f) summarized the changes in the relative density of p-NR1 immunoreactivity in the L1 (e) and S1 (f) spinal dorsal horn at 3 days and 7 days of colitis. Microphotographs were from 3 days of colitis and control animals. *p < 0.05 vs. control. Bar = 250 μm
BDNF regulated NR1 phosphorylation on Ser896 in vivo and in culture
BDNF generated by primary afferent neurons is able to release to the axonal terminals upon stimulation. The primary afferent neurons that innervate the distal colon also project to the spinal cord dorsal horn due to their pseudounipolar nature with two split branches, one going to the periphery and another going to spinal cord. During colitis, the level of BDNF was elevated in the DRG [9], specifically in the colonic afferent neurons [47]. Thus, we examined whether BDNF was able to elicit NR1 phosphorylation at Ser896. Using an acute system of spinal cord culture as described previously [5, 9], we found that incubation of the spinal cord slices with BDNF increased the phosphorylation level of NR1 at Ser896, which was effectively blocked by pre-treatment of the spinal slices with Trk inhibitor K252a (100 nM, Calbiochem) (Fig. 3), suggesting that NR1 phosphorylation at Ser896 by BDNF was specific and was mediated by BDNF high affinity receptor. We then examined the role of endogenous BDNF on NR1 phosphorylation at Ser896. In both L1 and S1 spinal cord, neutralization of BDNF action in vivo with specific BDNF antibody reversed colitis-induced NR1 phosphorylation (Fig. 4). We confirmed these findings in BDNF+/− rat (SAGE® Labs, Boyertown, PA) (Fig. 4).

BDNF induced NR1 phosphorylation in the spinal cord which was reduced by K252a. Western blot (a) showed that incubation of the acutely cultured spinal cord slices with BDNF (50 ng/mL) elicited an increase in the phosphorylation level of NR1 at Ser896 (b). Pre-treatment of the spinal cord slices with a TrkB inhibitor K252a reduced BDNF-elicited NR1 phosphorylation (c). *p < 0.05 vs. control or DMSO alone; #p < 0.05 vs. BDNF (+DMSO)

Up-regulation of NR1 phosphorylation in the spinal cord during colitis was regulated by endogenous BDNF. Western blot analysis of the L1 (a) and S1 (c) spinal cord from control rats, rats receiving TNBS or TNBS + BDNF antibody, and rats with partial deletion of BDNF (BDNF+/−) showed that blockade of BDNF action in vivo with BDNF neutralizing antibody or reduction of endogenous BDNF level by genetically engineering attenuated colitis-induced NR1 phosphorylation (b, d). *p < 0.05 vs. control; #p < 0.05 vs. TNBS
Signaling pathways in BDNF-induced NR1 phosphorylation in the spinal cord
We examined the involvement of three pathways, the MEK/ERK, PLCγ, and PI3K/Akt, in BDNF-induced NR1 phosphorylation at Ser896 in spinal slice culture. This was an acute system for quick initial screening of pathways. We used specific inhibitors: PD98059 to block MEK, U73122 to block PLCγ, and LY294002 to block PI3K. Since activation of PLCγ could facilitate DAG production leading to PKC activation, and NR1 Ser896 was considered to be phosphorylated by PKC [20], we also used bisindolylmaleimide I (BIM) to specifically block PKC in this system. Results (Fig. 5) showed that PD98059 (5 μM) did not have an effect on BDNF-induced NR1 phosphorylation; thus, the MEK/ERK pathway was not involved. The PLC inhibitor U73122 (10 μM) and PKC inhibitor BIM (1 μM) were able to attenuate BDNF-induced NR1 phosphorylation. The PI3K/Akt pathway was initially examined by using inhibitor LY294002 (5 μM) and then confirmed by another PI3K inhibitor wortmannin (0.5 μM). Both inhibitors blocked BDNF-induced NR1 phosphorylation at Ser896 (Fig. 5).

Signal transduction study of BDNF-induced NR1 phosphorylation in the spinal cord. The spinal cord slices were randomly separated into groups and pre-treated with a variety of specific inhibitors 30 min prior to BDNF stimulation. These inhibitors included PD98059 (PD) to block the MEK/ERK pathway, U73122 (U) to block the PLC pathway, bisindolylmaleimide I (BIM) to block PKC, LY294002 (LY), and wortmannin (WM) to block the PI3K/Akt pathway. Western blot (a) showed that PD had no effect on BDNF-induced NR1 phosphorylation while the rest of the inhibitors attenuated BDNF-elicited phosphorylation of NR1 at Ser896. Results were summarized in (b) indicating that the PLC/PKC and the PI3K/Akt, but not the MEK/ERK pathway was involved in BDNF-initiated NR1 phosphorylation. *p < 0.05 vs. control; #p < 0.05 vs. BDNF
NR1 phosphorylation at Ser896 in the spinal cord was mediated by the PI3K/Akt pathway in vivo in colitis
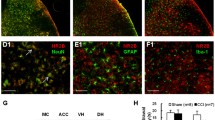
The involvement of the PI3K/Akt pathway in NR1 phosphorylation at Ser896 has not been reported previously. Thus, we confirmed our in vitro data (Fig. 5) with the in vivo system in animals induced for colitis. Double immunostaining (Fig. 6) showed that the p-NR1 (Fig. 6a, red stain) and p-Akt (Fig. 6b, green stain) had similar distribution pattern in the spinal cord (sections were L1 from TNBS 7 days). Higher magnification with ApoTome scan visualized the co-localization of p-NR1 and p-Akt in the spinal dorsal horn region (Fig. 6d–f, white arrows). These results suggested an association of Akt with NR1 phosphorylation in the spinal cord. We then injected the PI3K inhibitor LY294002 to colitic animals to block the PI3K/Akt activity in vivo (Fig. 7). We previously found that colitis increased the phosphorylation level of Akt in the L1 and S1 spinal cord at 7 days post-TNBS treatment [46]. Here, we found that LY294002 treatment reduced the Akt activity in the spinal cord during colitis examined by western blot (Fig. 7a, b showed S1 spinal cord; similar results were seen in L1 spinal cord, data not shown). Immunostaining showed that LY294002 treatment reduced p-Akt immunoreactivity in the spinal dorsal horn region during colitis (Fig. 7c, d showed L1 spinal cord; similar results were seen in S1 spinal cord, data not shown). LY294002 treatment also reduced colitis-induced NR1 phosphorylation at Ser896 (Fig. 7e, f).

Co-localization of phospho-Akt and phospho-NR1 in the spinal cord. Double immunostaining of the spinal cord from TNBS animals showed that p-NR1 (a, d, red cells) and p-Akt (b, e, green cells) had similar anatomic distribution which included the dorsal horn and deep laminae (a–c). ApoTome scan revealed that majority of p-NR1 (d, red cells) and p-Akt (e, green cells) were expressed in the same cells (f, cells indicated by white arrows). Some of the p-NR1 positive cells did not express p-Akt (d–f, yellow arrows). Also some of the p-Akt positive cells did not express p-NR1 (d–f, blue arrows). Figure showed samples of the L1 spinal cord from animals at 7 days of colitis. Bar = 200 μm in a–c; 15 μm in d–f

Inhibition of the PI3K in vivo with LY294002 blocked colitis-induced Akt activity and NR1 phosphorylation in the spinal cord. Animals receiving TNBS treatment (7 days) were divided into two groups: one receiving vehicle as control and one receiving LY294002 treatment. Animals treated with LY294002 had a reduced level of p-Akt in the spinal cord examined by western blot (a, b) and immunohistochemistry (c, d, circled areas are spinal dorsal horn), suggesting that inhibition of PI3K blocked Akt activity in vivo. Animals treated with LY294002 also had a reduced level of phospho-NR1 at Ser896 (e, f). Figure showed samples from S1 spinal cord. *p < 0.05. Bar = 250 μm
Discussion
Colitis-induced spinal central sensitization is attributable to posttranscriptional and posttranslational regulation in the spinal dorsal horn and is modulated by mediators generated in the primary afferent neurons. The NMDA receptor is one of the key molecules in regulating neuronal excitability and synaptic plasticity. The present study investigated the signaling pathways by which colonic inflammation facilitated phosphorylation of the NR1 subunit of the NMDA receptor in the spinal dorsal horn and identified that endogenous and exogenous BDNF were able to elicit NR1 phosphorylation at Ser896. In the spinal cord, colitis caused an increase in the level of NR1 Ser896 phosphorylation which was attenuated by inhibition of endogenous BDNF action with a specific BDNF antibody. The level of NR1 phospho-Ser896 was also reduced in BDNF+/− rats treated with TNBS. The signaling pathways that transduced BDNF signal in the spinal cord leading to NR1 phosphorylation at Ser896 included the PLCγ, PKC, and the PI3K/Akt pathways. Even though the ERK pathway was also activated in the spinal cord by BDNF during colitis [9], the activation of this pathway did not lead to NR1 phosphorylation at Ser896. The phosphorylation of NR1 at Ser896 was traditionally recognized as the PKC site and was phosphorylated by PKC [20]. The present study revealed an additional pathway, i.e., the PI3K/Akt pathway, in mediating NR1 Ser896 phosphorylation in the spinal cord. The PI3K/Akt pathway was reported to mediate substance P (SP)-regulated NR2B subunit of the NMDA receptor [4], but had no role in CGRP-induced NR1 phosphorylation at Ser897 [5]. Taken together, these results suggest that inflammation-induced spinal central sensitization characterized by site- and isoform-specific phosphorylation of the NMDA receptor is regulated by primary afferent release of neuropeptides and neurotransmitters such as BDNF that is examined in the present study. Specific signal transduction pathways are involved.
Visceral hypersensitivity is a highly complex entity that can occur due to hyperexcitability of the primary sensory afferents and dysregulation of spinal neurons (central sensitization) that modulate nociceptive transmission. Following noxious stimulation (e.g., colitis in the present study), excitatory neurotransmitters such as BDNF, substance P, and CGRP release centrally from the primary afferent neurons into the spinal dorsal horn [46, 48–50] where they can bind to their respective receptors and facilitate signal transduction [4, 9, 35, 46], ultimately resulting in pain hypersensitivity (central sensitization) [51]. The mechanism and contribution of neuropeptides to central sensitization has been well discussed by Seybold [52]. These neuropeptides are often stored in large dense core vesicles of unmyelinated (C) and small myelinated (Aδ) terminals, and their release are triggered by higher firing frequency-evoked intracellular Ca2+ concentration and/or by persistent stimuli [53–56]. Neuropeptides sometimes also co-store with small molecule transmitters such as glutamate which is released from both the large and small myelinated fibers that can synapse on NMDA receptor-containing neurons in the spinal cord in regulation of nociception [57, 58].
The biological responses to neuropeptide release in the spinal cord involve a series of changes in the intracellular components. Activation of kinases is a key process in receptor-mediated signal transduction. The present study characterized three major pathways that transduce the BDNF signaling in neurons: the MEK/ERK pathway, the PI3K/Akt pathway, and the PLCγ pathway. Among these three pathways, MEK/ERK pathway is activated by BDNF in the spinal cord during colitis [9]; however, it is not involved in BDNF-induced NR1 phosphorylation at Ser896 characterized in the present study. We have previously reported that the phosphorylation level of Akt is increased in the L1 and S1 spinal cord during colitis [46]. In the present study, the PI3K/Akt pathway also mediates BDNF- and colitis-induced NR1 phosphorylation at Ser896 in the spinal cord. This is an additional pathway to the widely accepted NR1 Ser896 phosphorylation mechanism by which PKC is involved [20]. Akt is traditionally considered as a survival factor targeting Bcl proteins, pro-caspase, and Forkhead [59, 60]. Several lines of recent research have suggested a critical role of the PI3K/Akt pathway in mediating sensory sensitization in response to peripheral inflammation or injury [4, 5, 61]. The activity of Akt is regulated by PI3K-facilitated formation of phosphatidylinositol (3,4,5)-trisphosphate (PIP3) which results in Akt trafficking and activation [62]. In the periphery, Akt is activated in the sensory neurons and regulates pain perception by activating TRPV1 receptor [61, 63]. In the spinal cord, the Akt activity can be regulated by BDNF, CGRP, and SP in a PI3K-dependent manner [4, 5, 46]. In terms of visceral inflammation, activation of the PI3K/Akt pathway is unable to regulate NR1 phosphorylation at Ser897, but mediates NR1 phosphorylation at Ser896 (the present study) in the spinal cord, suggesting that the PI3K/Akt pathway is important but not the sole pathway in visceral inflammation-induced NMDA receptor activation and spinal central sensitization.
The interrelationship of Akt activation and NMDA receptor activity has been demonstrated in several systems. A study in a formalin-induced hyperalgesia model shows that inhibition of the PI3K/Akt pathway blocks peripheral inflammation-induced phosphorylation of NR2 subunit of the NMDA receptor in the spinal cord [4]. In cerebellar granule, cell culture activation of Akt is also able to mediate forskolin-induced phosphorylation of Ser897 in the NR1 subunit [64]. In inflammatory and neuropathic pain models, the NR1 phosphorylation is enhanced in the spinal cord [65, 66], and in these studies, the antibodies used for NR1 phosphorylation are selective for either the Ser897 site alone or both the Ser897 and Ser896 sites together but not Ser896 alone. In our current study, we use an antibody that recognizes specifically Ser896 of the NR1 and find that this site is phosphorylated in vivo during colitis. We have previously tested that CGRP is unable to increase NR1 phosphorylation levels at Ser896 [5]. The present study demonstrates that p-NR1 Ser896 is regulated by BDNF-induced PLCγ and PKC cascade, consistent to the established findings [20, 67]. The present study also reveals that BDNF- and colitis-induced NR1 phosphorylation at Ser896 is regulated by the PI3K/Akt pathway. However, there is no biochemical evidence to show whether the PI3K and/or Akt can use Ser896 of NR1 as a phosphorylation substrate. Akt serves as a convergent point in signaling network and is activated by growth factors and G-protein coupled receptors and also cross talks with other signaling pathways in cells and tissues. In the complex PI3K/Akt signaling network, the mammalian target of rapamycin complex can promote the stability and activity of PKC [68]. In spinal slice culture, it takes 1 h for BDNF to stimulate NR1 phosphorylation, which implies that complicated signaling cross talk may exist within the spinal cord. Akt can be activated by a number of cytokines that are discovered in the spinal cord, such as the pro-inflammatory cytokine tumor necrosis factor (TNF) that is released from spinal glial cells [34, 69]. The involvement of cytokines in persistent pain has been well discussed recently [70]. These studies along with our current findings suggest a key modulatory role of the PI3K/Akt pathway in the generation and maintenance of visceral and somatic hypersensitivity and pain by being activated by a number of inflammatory and neuronal factors and regulating a number of ion channels.
Conclusion
The role of BDNF in mediating sensory hypersensitivity has been well recognized. The present study explores the molecular mechanisms and pathways that underlie the BDNF action in the spinal cord during colitis. BDNF in the spinal cord regulates NR1 phosphorylation which is mediated by specific signaling pathways. Especially, the PI3K/Akt pathway is found to be an additional pathway to PKC in regulating the NR1 phosphorylation at Ser896. The PI3K/Akt pathway is also important in the regulation of the NMDA receptor activity due to its ability in mediating NR2B subunit [4, 64]. The phosphorylation of these subunits is regulated specifically by diverse neuropeptides, such as BDNF, CGRP, and SP, converging on Akt and ultimately leads to NMDA receptor activation, which further mediates Ca2+ flux and modifies the strength and efficacy of synaptic transmission.
Abbreviations
- BDNF:
-
brain-derived neurotrophic factor
- BIM:
-
bisindolylmaleimide
- CGRP:
-
calcitonin gene-related peptide
- DRG:
-
dorsal root ganglia
- ERK:
-
extracellular signal-regulated kinase
- NMDA:
-
N-Methyl-D-aspartic acid
- NR1:
-
NMDA receptor 1
- PI3K:
-
phosphatidylinositol 3-kinase
- PKA:
-
protein kinase A
- PKC:
-
protein kinase C
- PLCγ:
-
phospholipase C-gamma
- TNBS:
-
tri-nitrobenzene sulfonic acid
References
Price DD, Zhou Q, Moshiree B, Robinson ME, Verne GN. Peripheral and central contributions to hyperalgesia in irritable bowel syndrome. J Pain. 2006;7(8):529–35.
Mayer EA. Commentary on peripheral and central contributions to hyperalgesia in irritable bowel syndrome. J Pain. 2006;7(8):539–41. discussion 542-3.
Agostini S, Eutamene H, Cartier C, Broccardo M, Improta G, Houdeau E, et al. Evidence of central and peripheral sensitization in a rat model of narcotic bowel-like syndrome. Gastroenterology. 2010;139(2):553–63. 563 e1-5.
Pezet S, Marchand F, D'Mello R, Grist J, Clark AK, Malcangio M, et al. Phosphatidylinositol 3-kinase is a key mediator of central sensitization in painful inflammatory conditions. J Neurosci. 2008;28(16):4261–70.
Kay JC, **a CM, Liu M, Shen S, Yu SJ, Chung C, et al. Endogenous PI3K/Akt and NMDAR act independently in the regulation of CREB activity in lumbosacral spinal cord in cystitis. Exp Neurol. 2013;250:366–75.
Burton MB, Gebhart GF. Effects of intracolonic acetic acid on responses to colorectal distension in the rat. Brain Res. 1995;672(1-2):77–82.
Coutinho SV, Meller ST, Gebhart GF. Intracolonic zymosan produces visceral hyperalgesia in the rat that is mediated by spinal NMDA and non-NMDA receptors. Brain Res. 1996;736(1-2):7–15.
Larsson MH, Rapp L, Lindstrom E. Effect of DSS-induced colitis on visceral sensitivity to colorectal distension in mice. Neurogastroenterol Motil. 2006;18(2):144–52.
Qiao LY, Gulick MA, Bowers J, Kuemmerle JF, Grider JR. Differential changes in brain-derived neurotrophic factor and extracellular signal-regulated kinase in rat primary afferent pathways with colitis. Neurogastroenterol Motil. 2008;20(8):928–38.
**a CM, Gulick MA, Yu SJ, Grider JR, Murthy KS, Kuemmerle JF, et al. Up-regulation of brain-derived neurotrophic factor in primary afferent pathway regulates colon-to-bladder cross-sensitization in rat. J Neuroinflammation. 2012;9:30.
Kohno T, Wang H, Amaya F, Brenner GJ, Cheng JK, Ji RR, et al. Bradykinin enhances AMPA and NMDA receptor activity in spinal cord dorsal horn neurons by activating multiple kinases to produce pain hypersensitivity. J Neurosci. 2008;28(17):4533–40.
van der Heide LP, Kamal A, Artola A, Gispen WH, Ramakers GM. Insulin modulates hippocampal activity-dependent synaptic plasticity in a N-methyl-d-aspartate receptor and phosphatidyl-inositol-3-kinase-dependent manner. J Neurochem. 2005;94(4):1158–66.
Xu ZZ, Zhang L, Liu T, Park JY, Berta T, Yang R, et al. Resolvins RvE1 and RvD1 attenuate inflammatory pain via central and peripheral actions. Nat Med. 2010;16(5):592–7. 1p following 597.
Iwata H, Takasusuki T, Yamaguchi S, Hori Y. NMDA receptor 2B subunit-mediated synaptic transmission in the superficial dorsal horn of peripheral nerve-injured neuropathic mice. Brain Res. 2007;1135(1):92–101.
Wang WT, Pan GQ, Zhang ZY, Suo ZW, Yang X, Hu XD. Ht31 peptide inhibited inflammatory pain by blocking NMDA receptor-mediated nociceptive transmission in spinal dorsal horn of mice. Neuropharmacology. 2015;89:290–7.
Ruscheweyh R, Wilder-Smith O, Drdla R, Liu X-G, Sandkühler J. Long-term potentiation in spinal nociceptive pathways as a novel target for pain therapy. Mol Pain. 2011;7:20.
Li F, Tsien JZ. Memory and the NMDA receptors. N Engl J Med. 2009;361(3):302–3.
Masu M, Nakajima Y, Moriyoshi K, Ishii T, Akazawa C, Nakanashi S. Molecular characterization of NMDA and metabotropic glutamate receptors. Ann N Y Acad Sci. 1993;707:153–64.
Mori H, Mishina M. Structure and function of the NMDA receptor channel. Neuropharmacology. 1995;34(10):1219–37.
Tingley WG, Ehlers MD, Kameyama K, Doherty C, Ptak JB, Riley CT, et al. Characterization of protein kinase A and protein kinase C phosphorylation of the N-methyl-D-aspartate receptor NR1 subunit using phosphorylation site-specific antibodies. J Biol Chem. 1997;272(8):5157–66.
Zou X, Lin Q, Willis WD. Role of protein kinase A in phosphorylation of NMDA receptor 1 subunits in dorsal horn and spinothalamic tract neurons after intradermal injection of capsaicin in rats. Neuroscience. 2002;115(3):775–86.
Brenner GJ, Ji RR, Shaffer S, Woolf CJ. Peripheral noxious stimulation induces phosphorylation of the NMDA receptor NR1 subunit at the PKC-dependent site, serine-896, in spinal cord dorsal horn neurons. European Journal of Neuroscience. 2004;20(2):375–84.
Obata K, Noguchi K. BDNF in sensory neurons and chronic pain. Neurosci Res. 2006;55(1):1–10.
Garraway SM, Petruska JC, Mendell LM. BDNF sensitizes the response of lamina II neurons to high threshold primary afferent inputs. Eur J Neurosci. 2003;18(9):2467–76.
Delafoy L, Gelot A, Ardid D, Eschalier A, Bertrand C, Doherty AM, et al. Interactive involvement of brain derived neurotrophic factor, nerve growth factor, and calcitonin gene related peptide in colonic hypersensitivity in the rat. Gut. 2006;55(7):940–5.
Groth R, Aanonsen L. Spinal brain-derived neurotrophic factor (BDNF) produces hyperalgesia in normal mice while antisense directed against either BDNF or trkB, prevent inflammation-induced hyperalgesia. Pain. 2002;100(1-2):171–81.
Matayoshi S, Jiang N, Katafuchi T, Koga K, Furue H, Yasaka T, et al. Actions of brain-derived neurotrophic factor on spinal nociceptive transmission during inflammation in the rat. J Physiol-London. 2005;569(2):685–95.
Wang LN, Yang JP, Ji FH, Zhan Y, ** XH, Xu QN, et al. Brain-derived neurotrophic factor modulates N-methyl-D-aspartate receptor activation in a rat model of cancer-induced bone pain. J Neurosci Res. 2012;90(6):1249–60.
Ng BK, Chen L, Mandemakers W, Cosgaya JM, Chan JR. Anterograde transport and secretion of brain-derived neurotrophic factor along sensory axons promote Schwann cell myelination. J Neurosci. 2007;27(28):7597–603.
Chen J, Winston JH, Sarna SK. Neurological and cellular regulation of visceral hypersensitivity induced by chronic stress and colonic inflammation in rats. Neuroscience. 2013;248:469–78.
Uchida H, Matsushita Y, Ueda H. Epigenetic regulation of BDNF expression in the primary sensory neurons after peripheral nerve injury: implications in the development of neuropathic pain. Neuroscience. 2013;240:147–54.
Cao XH, Chen SR, Li L, Pan HL. Nerve injury increases brain-derived neurotrophic factor levels to suppress BK channel activity in primary sensory neurons. J Neurochem. 2012;121(6):944–53.
Yoshii A, Constantine-Paton M. Postsynaptic BDNF-TrkB signaling in synapse maturation, plasticity, and disease. Dev Neurobiol. 2010;70(5):304–22.
Choi JI, Svensson CI, Koehrn FJ, Bhuskute A, Sorkin LS. Peripheral inflammation induces tumor necrosis factor dependent AMPA receptor trafficking and Akt phosphorylation in spinal cord in addition to pain behavior. Pain. 2010;149(2):243–53.
Xu Q, Fitzsimmons B, Steinauer J, O'Neill A, Newton AC, Hua XY, et al. Spinal phosphinositide 3-kinase-Akt-mammalian target of rapamycin signaling cascades in inflammation-induced hyperalgesia. J Neurosci. 2011;31(6):2113–24.
Cataldo G, Rajput S, Gupta K, Simone DA. Sensitization of nociceptive spinal neurons contributes to pain in a transgenic model of sickle cell disease. Pain. 2015;156(4):722–30.
He YQ, Chen Q, Ji L, Wang ZG, Bai ZH, Stephens Jr RL, et al. PKC gamma receptor mediates visceral nociception and hyperalgesia following exposure to PTSD-like stress in the spinal cord of rats. Mol Pain. 2013;9:35.
Melemedjian OK, Tillu DV, Asiedu MN, Mandell EK, Moy JK, Blute VM, et al. BDNF regulates atypical PKC at spinal synapses to initiate and maintain a centralized chronic pain state. Mol Pain. 2013;9:12.
Crozier RA, Bi C, Han YR, Plummer MR. BDNF modulation of NMDA receptors is activity dependent. J Neurophysiol. 2008;100(6):3264–74.
Geng SJ, Liao FF, Dang WH, Ding X, Liu XD, Cai J, et al. Contribution of the spinal cord BDNF to the development of neuropathic pain by activation of the NR2B-containing NMDA receptors in rats with spinal nerve ligation. Exp Neurol. 2010;222(2):256–66.
Ding X, Cai J, Li S, Liu XD, Wan Y, **ng GG. BDNF contributes to the development of neuropathic pain by induction of spinal long-term potentiation via SHP2 associated GluN2B-containing NMDA receptors activation in rats with spinal nerve ligation. Neurobiol Dis. 2015;73:428–51.
Kim JH, Roberts DS, Hu Y, Lau GC, Brooks-Kayal AR, Farb DH, et al. Brain-derived neurotrophic factor uses CREB and Egr3 to regulate NMDA receptor levels in cortical neurons. J Neurochem. 2012;120(2):210–9.
Slack SE, Thompson SW. Brain-derived neurotrophic factor induces NMDA receptor 1 phosphorylation in rat spinal cord. Neuroreport. 2002;13(15):1967–70.
Slack SE, Pezet S, McMahon SB, Thompson SW, Malcangio M. Brain-derived neurotrophic factor induces NMDA receptor subunit one phosphorylation via ERK and PKC in the rat spinal cord. Eur J Neurosci. 2004;20(7):1769–78.
**a CM, Colomb Jr DG, Akbarali HI, Qiao LY. Prolonged sympathetic innervation of sensory neurons in rat thoracolumbar dorsal root ganglia during chronic colitis. Neurogastroenterol Motil. 2011;23(8):801–e339.
Qiao LY, Grider JR. Colitis induces calcitonin gene-related peptide expression and Akt activation in rat primary afferent pathways. Exp Neurol. 2009;219(1):93–103.
Yu SJ, Grider JR, Gulick MA, **a CM, Shen S, Qiao LY. Up-regulation of brain-derived neurotrophic factor is regulated by extracellular signal-regulated protein kinase 5 and by nerve growth factor retrograde signaling in colonic afferent neurons in colitis. Exp Neurol. 2012;238(2):209–17.
Duggan AW, Hendry IA, Morton CR, Hutchison WD, Zhao ZQ. Cutaneous stimuli releasing immunoreactive substance-P in the dorsal horn of the cat. Brain Res. 1988;451(1-2):261–73.
Morton CR, Hutchison WD. Release of sensory neuropeptides in the spinal cord: studies with calcitonin gene-related peptide and galanin. Neuroscience. 1989;31(3):807–15.
Galeazza MT, Garry MG, Yost HJ, Strait KA, Hargreaves KM, Seybold VS. Plasticity in the synthesis and storage of substance-P and calcitonin-gene-related peptide in primary afferent neurons during peripheral inflammation. Neuroscience. 1995;66(2):443–58.
Choi JIL, Koehrn FJ, Sorkin LS. Carrageenan induced phosphorylation of Akt is dependent on neurokinin-1 expressing neurons in the superficial dorsal horn. Mol Pain. 2012;8:4.
Seybold VS. The role of peptides in central sensitization. Handb Exp Pharmacol. 2009;194:451–91.
Durham PL, Masterson CG. Two mechanisms involved in trigeminal CGRP release: implications for migraine treatment. Headache. 2013;53(1):67–80.
Meng JH, Ovsepian SV, Wang J, Pickering M, Sasse A, Aoki KR, et al. Activation of TRPV1 mediates calcitonin gene-related peptide release, which excites trigeminal sensory neurons and is attenuated by a retargeted botulinum toxin with anti-nociceptive potential. J Neurosci. 2009;29(15):4981–92.
Qin N, Neeper MP, Liu Y, Hutchinson TL, Lubin ML, Flores CM. TRPV2 is activated by cannabidiol and mediates CGRP release in cultured rat dorsal root ganglion neurons. J Neurosci. 2008;28(24):6231–8.
Schweitzer ES, Jeng CJ, TaoCheng JH. Selective localization and regulated release of calcitonin gene-related peptide from dense-core vesicles in engineered PC12 cells. J Neurosci Res. 1996;46(5):519–30.
De Biasi S, Rustioni A. Glutamate and substance P coexist in primary afferent terminals in the superficial laminae of spinal cord. Proc Natl Acad Sci U S A. 1988;85(20):7820–4.
Carozzi V, Marmiroli P, Cavaletti G. Focus on the role of glutamate in the pathology of the peripheral nervous system. CNS & Neurological Disorders-Drug Targets. 2008;7(4):348–60.
Amaravadi R, Thompson CB. The survival kinases Akt and Pim as potential pharmacological targets. Journal of Clinical Investigation. 2005;115(10):2618–24.
Manning BD, Cantley LC. AKT/PKB signaling: navigating downstream. Cell. 2007;129(7):1261–74.
Zhu W, Oxford GS. Phosphoinositide-3-kinase and mitogen activated protein kinase signaling pathways mediate acute NGF sensitization of TRPV1. Mol Cell Neurosci. 2007;34(4):689–700.
Toker A, Newton AC. Cellular signaling: pivoting around PDK-1. Cell. 2000;103(2):185–8.
Qiao LY, Yu SJ, Kay JC, **a CM. In vivo regulation of brain-derived neurotrophic factor in dorsal root ganglia is mediated by nerve growth factor-triggered Akt activation during cystitis. Plos One. 2013;8(11):e81547.
Llansola M, Sánchez-Pérez AM, Montoliu C, Felipo V. Modulation of NMDA receptor function by cyclic AMP in cerebellar neurones in culture. J Neurochem. 2004;91(3):591–9.
Gao X, Kim HK, Chung JM, Chung K. Enhancement of NMDA receptor phosphorylation of the spinal dorsal horn and nucleus gracilis neurons in neuropathic rats. Pain. 2005;116(1-2):62–72.
Zou X, Lin Q, Willis WD. Enhanced phosphorylation of NMDA receptor 1 subunits in spinal cord dorsal horn and spinothalamic tract neurons after intradermal injection of capsaicin in rats. J Neurosci. 2000;20(18):6989–97.
Michailidis IE, Helton TD, Petrou VI, Mirshahi T, Ehlers MD, Logothetis DE. Phosphatidylinositol-4,5-bisphosphate regulates NMDA receptor activity through alpha-actinin. J Neurosci. 2007;27(20):5523–32.
Fruman DA, Rommel C. PI3K and cancer: lessons, challenges and opportunities. Nat Rev Drug Discov. 2014;13(2):140–56.
Schafers M, Sorkin L. Effect of cytokines on neuronal excitability. Neurosci Lett. 2008;437(3):188–93.
Taves S, Berta T, Chen G, Ji R-R. Microglia and spinal cord synaptic plasticity in persistent pain. Neural Plast. 2013;2013, 753656.
Acknowledgements
This work was supported by grant NIH/NIDDK DK077917 to LYQ.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
ML, JCK, and LYQ designed and conducted most of the experiments. SS designed and conducted some of the experiments. ML and LYQ wrote most of the manuscript. All authors analyzed the data, revised the manuscript, and gave final approval for publication.
Additional file
Additional file 1:
Supplemental data. (PDF 18 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Liu, M., Kay, J.C., Shen, S. et al. Endogenous BDNF augments NMDA receptor phosphorylation in the spinal cord via PLCγ, PKC, and PI3K/Akt pathways during colitis. J Neuroinflammation 12, 151 (2015). https://doi.org/10.1186/s12974-015-0371-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12974-015-0371-z