
Abstract
Background
Caffeine exposure modifies the turnover of monoamine neurotransmitters, which play a role in several neuropsychiatric disorders. We conducted a Mendelian randomization study to investigate whether higher plasma caffeine levels are causally associated with the risk of anorexia nervosa, bipolar disorder, major depressive disorder (MDD), and schizophrenia.
Methods
Summary-level data on the neuropsychiatric disorders were obtained from large-scale genome-wide association studies (GWASs) of European ancestry participants (n = 72,517 to 807,553) and meta-analyzed with the corresponding data from the FinnGen study (n = 356,077). Summary-level data on plasma caffeine were extracted from a GWAS meta-analysis of 9876 European ancestry individuals. The Mendelian randomization analyses estimated the Wald ratio for each genetic variant and meta-analyzed the variant-specific estimates using multiplicative random effects meta-analysis.
Results
After correcting for multiple testing, genetically predicted higher plasma caffeine levels were associated with higher odds of anorexia nervosa (odds ratio [OR] = 1.124; 95% confidence interval [CI] = 1.024–1.238, pFDR = 0.039) and a lower odds of bipolar disorder (OR = 0.905, 95% CI = 0.827–0.929, pFDR = 0.041) and MDD (OR = 0.965, 95% CI = 0.937–0.995, pFDR = 0.039). Instrumented plasma caffeine levels were not associated with schizophrenia (OR = 0.986, 95% CI = 0.929–1.047, pFDR = 0.646).
Conclusions
These Mendelian randomization findings indicate that long-term higher plasma caffeine levels may lower the risk of bipolar disorder and MDD but increase the risk of anorexia nervosa. These results warrant further research to explore whether caffeine consumption, supplementation, or abstinence could render clinically relevant therapeutic or preventative psychiatric effects.
Similar content being viewed by others

Background
Caffeine is a widely consumed central nervous system stimulant found in coffee, tea, and cacao and is added to certain soft drinks, energy drinks, and analgesic drugs. The effects of caffeine are mainly mediated through the blockade of adenosine A1 and A2 receptors in the brain, which results in increased turnover of monoamine neurotransmitters, such as serotonin, dopamine, and noradrenaline [1]. Dysregulation of the monoamine neurotransmitter systems plays a role in several neuropsychiatric disorders, including anorexia nervosa [2], bipolar disorder [3], depression [4], and schizophrenia [5]. Furthermore, caffeine consumption is lower in people with depressive symptoms than in non-depressed individuals [6, 7], whereas caffeine consumption is higher in those with schizophrenia than in the general population [8]. However, whether caffeine consumption is causally associated with the risk of develo** neuropsychiatric disorders, or whether these disorders affect caffeine consumption, remains unestablished. Moreover, tolerance develops to some but not to all effects of caffeine [1]. Thus, whether long-term higher circulating caffeine levels affect the risk of neuropsychiatric disorders remains unclear.
The metabolism of caffeine is highly variable among individuals and depends in part on genetic variations in the activity of the enzyme cytochrome P450 1A2 (CYP1A2), which is responsible for over 90% of caffeine metabolism. For example, genetic variants in the CYP1A2 gene are associated with a lower paraxanthine to caffeine ratio (which reflects slower caffeine metabolism), higher plasma caffeine levels, and lower habitual caffeine consumption [9]. The latter association is likely related to the fact that persons with a genetic predisposition to slower caffeine metabolism require lower amounts of caffeine to reach the desired psychostimulant effect compared to individuals with faster caffeine metabolism.
To investigate whether long-term higher caffeine exposure causally associates with the risk of neuropsychiatric disorders, we conducted a Mendelian randomization (MR) study of the association between genetically predicted plasma caffeine levels and the risk of anorexia nervosa, bipolar disorder, major depressive disorder (MDD), and schizophrenia. Using genetic variants to instrument an exposure of interest (e.g., plasma caffeine) through MR reduces the biases that often limit observational studies, including reverse causation, as genetic variants cannot be modified by the outcome (such as neuropsychiatric disorders).
Methods
Study overview
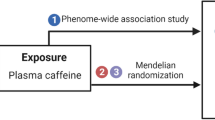
This study applied a two-sample MR design with publicly available summary-level data taken from meta-analyses of genome-wide association studies (GWASs) as well as the FinnGen study, which was not included in any of the GWAS meta-analyses (Fig. 1). Outcomes included in this study were anorexia nervosa, bipolar disorder, MDD, and schizophrenia. Brief information about the used data sources for plasma caffeine and outcomes are described below. More comprehensive details for each GWAS are described in the corresponding articles [9,10,11,12,13,14].

Study design overview (created with BioRender.com). AHR, aryl hydrocarbon receptor; CYP1A2, cytochrome P450 1A2 (CYP1A2); GWAS, genome-wide association study
Data sources
Plasma caffeine GWAS
Cornelis et al. performed a meta-analysis of summary statistics from six plasma caffeine (1,3,7-trimethylxanthine) GWASs comprising 9876 participants of mainly European ancestry, with mean ages between 47 and 71 years [9, 15]. Participants were asked to fast prior to measurements being taken, and each study standardized plasma caffeine measurements. Association estimates accounted for confounding by smoking status and, when applicable, age, sex, and principal components of ancestry.
Anorexia nervosa GWAS
Watson et al.’s GWAS was a meta-analysis from the Psychiatric Genetic Consortium (PGC) Eating Disorders Working Group, which contained 16,992 cases of anorexia nervosa and 55,525 controls from 33 studies—mostly of European ancestry individuals [14, 16]. Studies were asked to adjust their analyses for the first five principal components of ancestry. The linkage disequilibrium score regression intercept for the GWAS implied the presence of an insignificant residual population structure.
Bipolar disorder GWAS
This study is a meta-analysis of 41,917 cases of bipolar disorder and 371,549 controls from the PGC and five population-based cohort studies including the UK Biobank (UKB) [16, 17]. Samples were restricted to participants of European ancestry, and cases were diagnosed using the Diagnostic and Statistics Manual or International Classification of Diseases guidelines. The genomic control inflation factor was between 0.97 and 1.05 for the participating studies, implying minimal residual population structure.
MDD GWAS
Howard et al. meta-analyzed GWAS summary statistics of 170,756 MDD cases and 329,443 controls from the PGC and the UK Biobank [11]. All study samples largely comprised individuals of European ancestry. Cases were identified using a combination of self-report and medical records. The linkage disequilibrium (LD) score regression intercept for the GWAS summary statistics used is 1.02 (standard error = 0.01) implying the presence of insignificant residual population structure. GWAS summary statistics were extracted from the OpenGWAS platform using the ID: ieu-b-102 [18, 19].
Schizophrenia GWAS
Trubetskoy et al. meta-analyzed genetic data from 90 cohorts within the PGC [10, 16]. The resulting sample included 76,755 schizophrenia cases and 243,649 controls. Participating studies used genomic quality control to control for inflated test statistics and adjusted for at least four principal components of ancestry.
FinnGen study
The FinnGen study is a large (n = 356,077 in round 8) population biobank, based in Finland, described in detail elsewhere [20, 21]. GWAS summary data on the FinnGen cohort (round 8) includes 390 individuals with anorexia nervosa (FinnGen phenotype ID: R18 ANOREXIA), 6562 with bipolar disorder (F5 BIPO), 39,747 with MDD (F5 DEPRESSIO), and 6522 with schizophrenia (F5 SCHZPHR) [22]. Cases were identified from medical records [20, 21]. These GWASs adjusted for age, sex, the first 10 genetic principal components, and genoty** batch [12].
Mendelian randomization analysis
We selected single-nucleotide polymorphisms (SNPs) that were strongly associated (p < 5 × 10−5) with plasma caffeine levels and located within 100 kb of the CYP1A2 and aryl hydrocarbon receptor (AHR) gene regions as instrumental variables (GRCh37/hg19 assembly by Ensembl: 15:75041185-75048543 and 7:17338246-17385776, respectively). These genes were selected owing to their role in caffeine metabolism [9]. Variants in these gene regions have been used as instrumental variables for plasma caffeine in previous MR studies [23, 24]. The statistical significance threshold of p < 5 × 10−5 was selected to ensure that the used SNPs would be strong instruments for plasma caffeine, while accounting for gene region-wide (a Bonferroni correction for the 955 SNPs measured by Cornelis et al. within the two gene regions provides a p ~ < 5 × 10−5). SNPs were clumped with a r2 of 0.3 and 10,000 kb windows using the TwoSampleMR R package [25, 26]. We used the false discovery rate inverse quantile transformation (FIQT) Winner’s curse correction to account for Winner’s curse [27]. We harmonized data sources using TwoSampleMR and excluded palindromic SNPs, which could not be aligned based on their allele frequency.
We firstly meta-analyzed the SNP-outcome associations from each GWAS meta-analysis with the equivalent outcome data in the FinnGen study using an inverse variance weighted meta-analysis. Secondly, we conducted the MR analysis. The primary estimator was the Wald ratio, which was defined as the ratio of the SNP-outcome association to the SNP-exposure association. Wald ratios were combined using a multiplicative inverse variance-weighted (IVW) random effects model, using SNPs’ LD matrix estimated from the European subsample of the 1000 Genomes Project to account for correlations between variants [28]. This was implemented using the code for random effects IVW (accounting for correlations) by Burgess et al. [28]. A correlated variants IVW estimator was chosen over a more conventional (independent variant) IVW estimator because of the potential to improve the precision of MR estimates in a cis setting. We used the Benjamini–Hochberg procedure to correct for multiple testing. A false discovery rate (FDR) adjusted p-value < 0.05 was regarded as statistically significant. Odds ratios (OR) presented here represent the multiplicative increase in the odds for each standard deviation (SD) increase in plasma caffeine levels.
Heterogeneity tests have been proposed as sensitivity analyses for pleiotropy in MR studies [29]. Because these methods have not been extended to cis settings with correlated variants, we first explore the robustness of outlier SNPs using leave-one-out analyses and then explore heterogeneity between the AHR and CYP1A2 genes. We additionally explore the heterogeneity in the MR estimates between data sources and replicate our analysis using only the lead SNP from each gene region (rs4410790 for AHR and rs2472297 for CYP1A2). To the best of our knowledge, there is no sample overlap between the exposure and outcome GWASs.
Results
After clum** and harmonization, we included 7 SNPs as instrumental variables for plasma caffeine in all analyses except for MDD, where 8 SNPs were included (Additional file 1: Tables S1-S3). The discrepancy in the number of SNPs is due to the different coverages of SNPs measured in the GWASs used. After accounting for LD among SNPs, these genetic instruments had an average F statistic of 44 and 25, respectively. This is consistent with approximately 2–4% bias in the MR estimates due to the use of weak instruments.
Our MR analysis indicated that each SD increase in genetically predicted plasma caffeine was associated with a 1.124 (95% confidence interval [CI] = 1.024 to 1.238, pFDR = 0.039) fold higher odds of anorexia nervosa (Fig. 2 and Additional file 1: Fig. S1). In contrast, there were inverse associations of genetically predicted plasma caffeine levels with bipolar disorder (OR = 0.905, 95% CI = 0.827 to 0.929, pFDR = 0.041) and MDD (OR = 0.965, 95% CI = 0.937 to 0.995, pFDR = 0.039). There was no evidence of an association with schizophrenia (OR = 0.986, 95% CI = 0.929 to 1.047, pFDR = 0.646).

Forest plot of two-sample MR estimates. “OR” represents the effect of a standard deviation increase in genetically predicted plasma caffeine levels on the odds of each outcome. AHR, aryl hydrocarbon receptor; CI, confidence interval; CYP1A2, cytochrome P450 1A2 (CYP1A2); OR, odds ratio
Our leave-one-out analyses (Additional file 1: Fig. S2) found that exclusion of the variant rs12903896 resulted in deflation towards the null for the CYP1A2 anorexia estimate, but resulted in more extreme estimates, further away from the null, for the other outcomes. This effect was much less pronounced on the combined CYP1A2 and AHR estimates. All the AHR estimates were broadly consistent after the exclusion of each AHR SNP in the leave-one-out sensitivity analysis. The results of the analysis including only the lead variants for each gene (rs4410790 for AHR and rs2472297 for CYP1A2) are similar to those in our primary analysis (Fig. 3), other than being less precise. Together, these imply that rs12903896 has not biased our estimates.

Forest plot of two-sample Mendelian randomization estimates using lead variants (rs4410790 for AHR and rs2472297 for CYP1A2) only. “OR” represents the effect of a standard deviation increase in genetically predicted plasma caffeine levels on the odds of each outcome. AHR, aryl hydrocarbon receptor; CI, confidence interval; CYP1A2, cytochrome P450 1A2 (CYP1A2); OR, odds ratio
We did not find strong evidence of heterogeneity between AHR and CYP1A2 estimates for anorexia nervosa (I2 = 0%), bipolar disorder (I2 = 0%), MDD (I2 = 41%), or schizophrenia (I2 = 0%). We also did not find evidence of heterogeneity between the two data sources for anorexia nervosa, bipolar disorder, or MDD (Fig. 4). However, there was evidence of heterogeneity for schizophrenia (I2 = 84%, p = 0.01), although an additive random effects meta-analysis of the MR estimates from the PGC and FinnGen still did not find evidence of an effect (95% CI: 0.72 to 1.14).

Heterogeneity in Mendelian randomization estimates from the two different data sources. “OR” represents the effect of a standard deviation increase in genetically predicted plasma caffeine levels on the odds of each outcome. PGC, Psychiatric Genetic Consortia; UKB, UK Biobank; CI, confidence interval; logOR, log odds ratio; OR, odds ratio
Discussion
This two-sample MR study found that genetically predicted long-term higher plasma caffeine levels were associated with a higher risk of anorexia nervosa, but a lower risk of bipolar disorder and MDD. We found no significant associations of genetically predicted plasma caffeine levels with the risk of schizophrenia.
The link between dietary caffeine intake and depressive symptoms has been reported in several observational, mainly cross-sectional studies, which were by nature of their design unable to investigate the direction of the association. A recent meta-analysis of observational studies reported that high versus low caffeine intake was associated with an overall 14% lower risk of future depressive symptoms in two cohort studies and an amalgamated 13% lower odds of depressive symptoms across 10 cross-sectional studies [7]. Our investigation confirms these findings and extends the evidence that higher plasma caffeine may play a causal role in reducing the risk of MDD as well as bipolar disorder for which epidemiological data on caffeine are scarce.
Caffeine may through adenosine antagonism play a role in anorexia nervosa as altered serotonergic and dopaminergic function is commonly observed in those suffering from this disorder [2]. Although anorexia nervosa patients often present with co-morbid psychiatric disorders such as MDD, the present MR study found that genetically predicted higher plasma caffeine was associated with an increased rather than decreased risk of anorexia nervosa. Since previous MR evidence is indicative of an association between higher plasma caffeine and lower body mass index [23], the observed effect may be a result of the more general association of plasma caffeine with weight loss. This novel finding warrants confirmation by further studies.
Although a positive association between caffeine intake and prevalent schizophrenia has been reported [8], we did not observe any association between genetically predicted plasma caffeine levels and schizophrenia risk. Thus, our finding suggests that caffeine is not a strong causal risk factor for this disorder and that findings in traditional epidemiological studies may arise spuriously due to residual confounding or reverse causation.
Summary data MR estimates should be interpreted as the average lifetime effect of greater exposure to caffeine [30]. However, the incidence of many mental disorders is not uniformly distributed across the lifespan. For example, anorexia nervosa is typically developed in adolescence or early adulthood. Future research may consider using longitudinal/individual-level data to explore whether this has implications for the presence of time-varying effects.
Our study has several strengths. The MR design reduces the risk of reverse causation bias and confounding by environmental factors. We further strengthen the validity of our findings by biologically plausible genetic instruments from genes known to affect caffeine metabolism and plasma caffeine levels. A priori gene selection additionally reduces the risk of confounding by pleiotropy. Finally, the use of large-scale GWAS data within a two-sample MR framework increased the statistical power of our analyses.
Nevertheless, despite the use of the largest publicly available GWAS data for the outcomes, we cannot exclude the possibility that we might have overlooked weak associations between plasma caffeine and the studied disorders. Furthermore, the possibility of genetic confounding by metabolites other than caffeine that are metabolized by the CYP1A2 enzyme cannot be ruled out. Although the use of a cis design should provide greater robustness through establishing the biological mechanism linking SNPs to the exposure [31], a limitation of it is that many standard MR sensitivity analyses cannot be used in settings with correlated variants. A further limitation is that the exposure GWAS adjusted for smoking status. Although adjustment for heritable covariates can create collider bias [32], the outcome associations were not adjusted for this variable. Therefore, while this adjustment may lead to bias, it will not affect the significance of MR estimates (as significance depends on the variant-outcome associations), and so inferences from our analyses should be valid [33, 34]. In addition, the use of summary-level data meant that dose–response relations of the effect of caffeine on psychiatric disorders could not be investigated. Finally, our MR analyses largely included data collected from individuals of European descent, and therefore, our findings may not be transferable to populations of non-European descent.
Conclusions
This MR investigation provides evidence of possible causal relationships of long-term higher plasma caffeine levels with a higher risk of anorexia nervosa and a reduced risk of bipolar disorder and MDD. These results warrant further research to explore whether caffeine consumption, supplementation, or abstinence could render clinically relevant therapeutic or preventative psychiatric effects.
Availability of data and materials
All data analyzed during the current study are available in Additional file 1: Tables S1 to S4. The R code is available at https://github.com/bar-woolf/applied-MR-code/blob/main/caff%20psyc.R. This study was not pre-registered.
Abbreviations
- AHR:
-
Aryl hydrocarbon receptor
- CI:
-
Confidence interval
- CYP1A2:
-
Cytochrome P450 1A2
- FDR:
-
False discovery rate
- GWAS:
-
Genome-wide association study
- IVW:
-
Inverse variance-weighted
- MDD:
-
Major depressive disorder
- MR:
-
Mendelian randomization
- OR:
-
Odds ratio
- PGC:
-
Psychiatric Genetic Consortium
- SNP:
-
Single-nucleotide polymorphism
References
Fredholm BB, Bättig K, Holmén J, Nehlig A, Zvartau EE. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev. 1999;51(1):83–133.
Bosanac P, Norman T, Burrows G, Beumont P. Serotonergic and dopaminergic systems in anorexia nervosa: a role for atypical antipsychotics? Aust N Z J Psychiatry. 2005;39(3):146–53.
Miklowitz DJ, Johnson SL. The psychopathology and treatment of bipolar disorder. Annu Rev Clin Psychol. 2006;2:199–235.
Heninger GR, Delgado PL, Charney DS. The revised monoamine theory of depression: a modulatory role for monoamines, based on new findings from monoamine depletion experiments in humans. Pharmacopsychiatry. 1996;29(1):2–11.
Brisch R, Saniotis A, Wolf R, Bielau H, Bernstein HG, Steiner J, et al. The role of dopamine in schizophrenia from a neurobiological and evolutionary perspective: old fashioned, but still in vogue. Front Psychiatry. 2014;5:47.
Bao J, Li P, Guo Y, Zheng Y, Smolinski M, He J. Caffeine is negatively associated with depression in patients aged 20 and older. Front Psychiatry. 2022;13:1037579.
Torabynasab K, Shahinfar H, Payandeh N, Jazayeri S. Association between dietary caffeine, coffee, and tea consumption and depressive symptoms in adults: a systematic review and dose-response meta-analysis of observational studies. Front Nutr. 2023;10:1051444.
Topyurek M, Tibbo P, Núñez C, Stephan-Otto C, Good K. Caffeine effects and schizophrenia: is there a need for more research? Schizophr Res. 2019;211:34–5.
Cornelis MC, Kacprowski T, Menni C, Gustafsson S, Pivin E, Adamski J, et al. Genome-wide association study of caffeine metabolites provides new insights to caffeine metabolism and dietary caffeine-consumption behavior. Hum Mol Genet. 2016;25(24):5472–82.
Trubetskoy V, Panagiotaropoulou G, Awasthi S, Braun A, Kraft J, Skarabis N, et al. Map** genomic loci implicates genes and synaptic biology in schizophrenia. Nature. 2022;604(7906):502–8.
Howard DM, Adams MJ, Shirali M, Clarke TK, Marioni RE, Davies G, et al. Genome-wide association study of depression phenotypes in UK Biobank identifies variants in excitatory synaptic pathways. Nat Commun. 2018;16(9):1470.
Risteys FinnGen R6 - I9_CVD. https://r6.risteys.finngen.fi/phenocode/I9_CVD. [cited 2022 Apr 6].
Watanabe K, Jansen PR, Savage JE, Nandakumar P, Wang X, Hinds DA, et al. Genome-wide meta-analysis of insomnia prioritizes genes associated with metabolic and psychiatric pathways. Nat Genet. 2022;54(8):1125–32.
Watson HJ, Yilmaz Z, Thornton LM, Hübel C, Coleman JR, Gaspar HA, et al. Genome-wide association study identifies eight risk loci and implicates metabo-psychiatric origins for anorexia nervosa. Nat Genet. 2019;51(8):1207–14.
Cornelis MC. caff_share.gz (v1.0.0). DigitalHub. Galter Health Sciences Library. 2016. https://doi.org/10.18131/G3860C.
Download Results – PGC. https://pgc.unc.edu/for-researchers/download-results/. [cited 2023 Jul 26].
Mullins N, Forstner AJ, O’Connell KS, Coombes B, Coleman JRI, Qiao Z, et al. Genome-wide association study of more than 40,000 bipolar disorder cases provides new insights into the underlying biology. Nat Genet. 2021;53(6):817–29.
Elsworth B, Lyon M, Alexander T, Liu Y, Matthews P, Hallett J, et al. The MRC IEU OpenGWAS data infrastructure. bioRxiv. 2020.08.10.244293.
Trait: Major depression - IEU OpenGWAS project https://gwas.mrcieu.ac.uk/datasets/ieu-b-102/. [cited 2023 Jul 26].
FinnGen. FinnGen Documentation of R5 release. 2021. https://finngen.gitbook.io/documentation/v/r5/.
Kurki MI, Karjalainen J, Palta P, Sipilä TP, Kristiansson K, Donner K, et al. FinnGen: unique genetic insights from combining isolated population and national health register data. medRxiv. 2022. 2022.03.03.22271360.
Access results | FinnGen https://www.finngen.fi/en/access_results [cited 2023 Jul 26].
Larsson SC, Woolf B, Gill D. Appraisal of the causal effect of plasma caffeine on adiposity, type 2 diabetes, and cardiovascular disease: two sample mendelian randomisation study. BMJ Med. 2023;2(1):1–8.
Larsson SC, Woolf B, Gill D. Plasma caffeine levels and risk of Alzheimer’s disease and Parkinson’s disease: Mendelian randomization study. Nutrients. 2022;14(9):1697.
R Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computin. 2021. https://www.R-project.org/.
Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, et al. The MR-Base platform supports systematic causal inference across the human phenome. eLife. 2018;7:e34408.
Bigdeli TB, Lee D, Riley BP, Vladimirov V, Fanous AH, Kendler KS, et al. FIQT: a simple, powerful method to accurately estimate effect sizes in genome scans. bioRxiv. 2015.019299.
Burgess S, Zuber V, Valdes-Marquez E, Sun BB, Hopewell JC. Mendelian randomization with fine-mapped genetic data: choosing from large numbers of correlated instrumental variables. Genet Epidemiol. 2017;41(8):714–25.
Bowden J, Hemani G, Davey SG. Invited commentary: detecting individual and global horizontal pleiotropy in Mendelian randomization—a job for the humble heterogeneity statistic? Am J Epidemiol. 2018;187(12):2681–5.
Labrecque JA, Swanson SA. Interpretation and potential biases of Mendelian randomization estimates with time-varying exposures. Am J Epidemiol. 2019;188(1):231–8.
Woolf B, Cronjé HT, Zagkos L, Larsson SC, Gill D, Burgess S. Comparison of caffeine consumption behavior with plasma caffeine levels as exposures in drug-target Mendelian randomization and implications for interpreting effects on obesity. medRxiv. 2023.05.30.23290752.
Hartwig FP, Tilling K, Davey Smith G, Lawlor DA, Borges MC. Bias in two-sample Mendelian randomization when using heritable covariable-adjusted summary associations. Int J Epidemiol. 2021;50(5):1639–50.
Lawlor DA, Harbord RM, Sterne JAC, Timpson N, Davey SG. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. 2008;27(8):1133–63.
Didelez V, Sheehan N. Mendelian randomization as an instrumental variable approach to causal inference. Stat Methods Med Res. 2007;16(4):309–30.
Acknowledgements
The authors thank the investigators of the GWAS publications on anorexia nervosa, bipolar disorder, MDD, and schizophrenia as well as the FinnGen study for making their data publicly available.
Funding
Open access funding provided by Uppsala University. B.W. is funded by an Economic and Social Research Council (ESRC) South West Doctoral Training Partnership (SWDTP) 1 + 3 PhD Studentship Award (ES/P000630/1) and a member of the Medical Research Council (MRC) Integrative Epidemiology Unit at the University of Bristol (MC_UU_00011/7). H.T.C. is supported by the Novo Nordic Foundation Challenge Programme: Harnessing the Power of Big Data to Address the Societal Challenge of Aging (NNF17OC0027812). D.G. is supported by the British Heart Foundation Centre of Research Excellence (RE/18/4/34215) at Imperial College. S.C.L. is supported by the Swedish Research Council for Health, Working Life and Welfare (Forte, 2018–00123); Swedish Heart Lung Foundation (Hjärt-Lungfonden, 20210351); the Swedish Cancer Society (Cancerfonden) and Swedish Research Council (Vetenskapsrådet, 2019–00977). S.B. is supported by the Wellcome Trust (225790/Z/22/Z) and the United Kingdom Research and Innovation Medical Research Council (MC_UU_00002/7). This research was supported by the National Institute for Health Research Cambridge Biomedical Research Centre (NIHR203312). The views expressed are those of the authors and not necessarily those of the National Institute for Health Research or the Department of Health and Social Care.
Author information
Authors and Affiliations
Contributions
B.W. performed the statistical analysis. B.W., H.T.C., and S.C.L. wrote the first draft of the manuscript. B.W., H.T.C., L.Z., S.B., D.G., and S.C.L. contributed to the design of the study and edited the manuscript for intellectual content. All authors read and approved the final manuscript
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Studies included in the published GWAS meta-analyses and the FinnGen study were approved by the relevant ethical review authority, and voluntary informed consent was obtained from participants. This study only analyzed publicly available summary-level statistical data. Therefore, no new ethical review board approval was required.
Consent for publication
Not applicable.
Competing interests
DG is employed part-time by Novo Nordisk. The other authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table S1. Exposure related genome-wide association study summary data taken from Cornelis et al. Table S2. Summary-level data from FinnGen. Table S3. Genome-wide association study summary statistics taken from other data sources. Table S4. Meta-analyzed variant-outcome associations and Mendelian randomization Wald ratios. Fig. S1. X-Y scatter graphs of Mendelian randomization results. Fig. S2. Leave-one-out analyses.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Woolf, B., Cronjé, H.T., Zagkos, L. et al. Appraising the causal relationship between plasma caffeine levels and neuropsychiatric disorders through Mendelian randomization. BMC Med 21, 296 (2023). https://doi.org/10.1186/s12916-023-03008-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12916-023-03008-0