
Abstract
Background
Adventitious roots (ARs) represent an important organ system for water and nutrient uptake in lotus plants because of degeneration of the principal root. The WUSCHEL-related homeobox (WOX) gene regulates plant development and growth by affecting the expression of several other genes. In this study, three WOX genes, NnWOX1-1, NnWOX4-3, and NnWOX5-1, were isolated and their functions were assessed in Arabidopsis plants.
Results
The full lengths of NnWOX1-1, NnWOX4-3, and NnWOX5-1 were 1038, 645, and 558 bp, encoding 362, 214, and 185 amino acid residues, respectively. Phylogenetic analysis classified NnWOX1-1 and NnWOX4-3 encoding proteins into one group, and NnWOX5-1 and MnWOX5 encoding proteins exhibited strong genetic relationships. The three genes were induced by sucrose and indoleacetic acid (IAA) and exhibited organ-specific expression characteristics. In addition to improving root growth and salt tolerance, NnWOX1-1 and NnWOX4-3 promoted stem development in transgenic Arabidopsis plants. A total of 751, 594, and 541 genes, including 19, 19, and 13 respective genes related to ethylene and IAA metabolism and responses, were enhanced in NnWOX1-1, NnWOX4-3, and NnWOX5-1 transgenic plants, respectively. Further analysis showed that ethylene production rates in transgenic plants increased, whereas IAA, peroxidase, and lignin content did not significantly change. Exogenous application of ethephon on lotus seedlings promoted AR formation and dramatically increased the fresh and dry weights of the plants.
Conclusions
NnWOX1-1, NnWOX4-3, and NnWOX5-1 influence root formation, stem development, and stress adaptation in transgenic Arabidopsis plants by affecting the transcription of multiple genes. Among these, changes in gene expression involving ethylene metabolism and responses likely critically affect the development of Arabidopsis plants. In addition, ethylene may represent an important factor affecting AR formation in lotus seedlings.
Similar content being viewed by others


Background
Lotus is a member of the Nymphaeaceae family [1, 2] and contains three subgroups (lotus root, seed lotus, and flowering lotus) according to usage. Lotus root is an essential vegetable in southern China [3], the region with the highest cultivation area of the aquatic vegetable within the country. Lotus root originated in China and its diverse products include lotus tea, lotus drinks, glutinous rice lotus, and salt lotus. These products are exported to Japan, South Korea, Europe, and the United States, resulting in substantial economic benefits for local farmers. Therefore, the planting area of lotus root has increased in open-air or protected facilities. In addition, lotus root can be used as an ingredient in Chinese herbal medicine [4], which is beneficial for human health. Generally, the storage organ is formed underground, and adventitious roots (ARs) are necessary for water and nutrient uptake owing to degeneration of the principal root. The development of ARs directly affects the yield and quality of lotus.
The ARs, usually derived from the root primordia, are always located in the hypocotyl, stem, and leaf, and strongly influence plant fixation as well as water and nutrient absorption [5, 6]. Therefore, ARs play an important role in plant growth, especially in those plants where the principal root has degenerated. Plant ARs are classified as a secondary root system, and three biological developmental processes that include the induction, initiation, and expression stages are observed during AR formation [7, 8]. In the first biological process, there is a transition of cell function, and undifferentiated cells develop into meristematic cells, which can further develop into ARs. The secondary biological stage is the primordial-established stage, and meristematic cells differentiate into primordial ARs [9]. In the third stage, the primordial ARs continually develop and break through the stem or leaf epidermis [10].
The formation of ARs is strictly regulated by genetic and environmental factors [11, 12]. Plant hormones are involved throughout the developmental stages of root formation [13]. Ethylene and indoleacetic acid (IAA) are considered the “promoters” of these three biological stages [14, 15]. In rice, ethylene affects AR formation by inducing the death of epidermal cells at the site of AR emergence [14]. Tomato seedlings treated with 1-aminocyclopropane-1-carboxylic acid (ACC, an intermediate metabolite in ethylene synthesis) contain more ARs than control plants [16]. IAA is also a critical factor that regulates AR formation in the plant kingdom [17]. Increasing the endogenous IAA content induces cell differentiation to form ARs [18], and anything that affects IAA synthesis and transport can alter the development of the lateral roots [19, 20]. The crosstalk between ethylene and IAA in regulating root formation shows that the AR formation process is highly complex and many physiological and biological metabolic processes are involved [21]. The formation of lotus ARs occurs in the hypocotyls of seedlings and the internodes of rhizomes [22]. Like in other plants, the formation of ARs in lotus is regulated by ethylene and IAA. Low concentrations of IAA promote cell differentiation, whereas high concentrations of exogenous IAA have the inverse effect [23]. Light and photosynthetic products also greatly influence AR formation [24, 25]. Lignin is closely related to AR formation in lotus seedlings. Increasing lignin content by overexpressing NnLAC17 leads to a decrease in root number in transgenic Arabidopsis plants [26], suggesting that the development of ARs in lotus plants involves a complex regulatory network. Additionally, abscisic acid (ABA) and cytokinins are involved in AR formation [27]. Gene regulation is a genetic factor that affects AR formation. Therefore, understanding the characteristics of gene expression is an effective method to monitor AR development. Several genes participate in AR development. The puroindoline (PIN) and AUX genes encode proteins that are influx and efflux carriers for IAA transport, which influence lateral root formation [28]. PIN is highly expressed in primordial organs [29]. Some genes with a lateral organ boundaries (LOB) domain are induced by ethylene and IAA and their expression is necessary for root primordium development. For example, the protein encoded by ADP-ribosylation factor-like GTPase 1 (ARL1), with its LOB domain, strongly influences cell dedifferentiation at the induction stage of ARs [30].
The WUSCHEL-related homeobox (WOX) gene family consists of several homeodomain transcription factors [31]. The members of the WOX family can be classified into three clades: ancient, intermediate, and WUSCHEL (WUS) [32,33,34]. This provides evidence that WOX influences stress adaptation and root formation in plants [35,36,37,38]. Some members of the WOX family regulate root development in the initial stage, which is triggered by auxins [39,40,41]. A similar role was found for WOX11, which regulates root architecture in Arabidopsis [42]. The overexpression of poplar WOX5a can compensate for a loss of function mutation in WOX5 in the Arabidopsis endogen [43]. Liu et al. [44] observed that several Populus tomentosa WOXs have different expression profiles, and their functions are the same for AR regeneration, thereby suggesting that WOXs are required for AR development.
In a previous study, we discovered that sucrose and IAA affected AR formation in lotus seedlings. At the same time, we also found that there was a crosstalk between the IAA and ethylene signaling pathways during AR formation [23, 25]. Based on transcriptome data, three transcription factors, NnWOX1-1, NnWOX4-3, and NnWOX5-1, were upregulated after IAA and sucrose treatment. Therefore, we assumed there were close connections between these three genes and hormones during AR development. In the present study, these three genes were cloned, and their expression profiles and functions were analyzed to further confirm the roles of these genes in AR development. In addition, a possible regulatory pathway of NnWOX1-1, NnWOX4-3, and NnWOX5-1 was explored.
Results
Cloning of NnWOX1-1, NnWOX4-3, and NnWOX5-1
Three WOXs were cloned via reverse transcription-polymerase chain reaction (RT-PCR). We observed that the full lengths of these three genes were 1038, 645, and 558 bp and encoded 345, 214, and 186 amino acid residues, respectively (Additional Table 1). After comparing with data from the National Center for Biotechnology Information (NCBI) database, it was observed that the three genes contained domains that are conserved in the WOX family from Arabidopsis, rice, corn, and peanut (Additional Fig. 1). Therefore, these genes were designated as NnWOX1-1, NnWOX4-3, and NnWOX5-1. The proteins encoded by NnWOX1-1, NnWOX4-3, and NnWOX5-1 had conserved regions, although little homology was found in these sequences (Fig. 1a). Further analysis revealed that the NnWOX1-1- and NnWOX4-3-encoded proteins each contained a homeobox domain, whereas the NnWOX5-1-encoded protein contained a superfamily homeodomain (Fig. 1b). The WOX1-, WOX4-, and WOX5-encoded proteins were subdivided into 14 groups. The NnWOX1-1- and NnWOX4-3-encoded proteins displayed distant relationships with others species and were classified into a single subgroup. The NnWOX5-1-encoded protein was closely related to that of MnWOX5 and was classified into another subgroup (Fig. 1c). Ten motifs were identified in the NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins, which contained eight, five, and seven motifs, respectively. Motifs 1, 2, and 3 were observed in all three proteins, whereas motifs 5 and 7 were observed only in the NnWOX1-1-encoded protein (Fig. 2a). Chromosomal localization was also performed; NnWOX1-1, NnWOX4-3, and NnWOX5-1 were located at NW-010729164.1, NW-010729076.1, and NW-010729104.1, respectively (Fig. 2b).

Comparison and phylogenetic tree analysis of Nnwox1-1, Nnwox4-3, and Nnwox5-1. a Comparison of NnWOX1-1, NnWOX4-3, and NnWOX5-1 with amino acid sequences. The red box represents the homologous region of the three genes. b Domain analysis of NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins, and boxes of different colors represent conserved regions. c Phylogenetic tree analysis of NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins with WOX-encoded proteins of other species. Fourteen groups with different colors were detected. The red triangle represents the positions of the three proteins in the phylogenetic tree

Conserved motif and chromosomal localization analysis of NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins in the whole lotus genome. a Distribution of conserved motifs in NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins. Boxes of different colors represent the ten putative motifs, and the boxes with the same color represent the same motif in the structure of these three genes. b Chromosomal localization analysis of NnWOX1-1, NnWOX4-3, and NnWOX5-1
Expression analysis of NnWOX1-1, NnWOX4-3, and NnWOX5-1
Reverse transcription quantitative PCR (RT-qPCR) was used to monitor the expression profiles of NnWOX1-1, NnWOX4-3, and NnWOX5-1 in lotus seedlings treated with sucrose and IAA. The expression of these three genes increased in response to sucrose treatment. NnWOX1-1 exhibited an enhanced mRNA level after 2 d, and NnWOX4-3 and NnWOX5-1 expression increased after 4 d of treatment. The same expression pattern was observed in response to IAA treatment. The transcriptional levels of NnWOX1-1, NnWOX4-3, and NnWOX5-1 were enhanced 2 d after IAA treatment and exhibited an increasing tendency within 6 d. Tissue-specific expression analysis showed that these three genes had different expression patterns in different organs. NnWOX1-1 mRNA levels were higher in the stem than in the ARs, leaves, and flowers. Increased NnWOX4-3 expression was observed in leaves, although the gene was expressed in other organs. The transcriptional level of NnWOX5-1 was higher in ARs than in leaves, stems, and flowers (Fig. 3).

Expression patterns of NnWOX1-1, NnWOX4-3, and NnWOX5-1 with different treatments and in different organs, as determined by RT-qPCR. a Expression analysis after sucrose treatment. b Identification of gene expression in lotus seedlings treated with IAA. c Organ-specific expression analysis in lotus plant roots, stems, leaves, and flowers. The mean expression values were calculated from three independent biological replicates, and “*” indicates values that are significantly different compared with control plants (P < 0.05)
Functional analysis of genes for root development
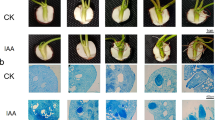
pSN1301:NnWOX1-1, pSN1301:NnWOX4-3, and pSN1301:NnWOX5-1 were constructed and transferred to Arabidopsis plants to identify the gene functions. “Positive” plants were identified by PCR (Additional Fig. 2). Seeds of the T2 generation were sown on base material (V:V soil: vermiculite = 1:1) and Murashige and Skoog (MS) culture medium. Transgenic NnWOX1-1 plants had longer stems than wild-type plants. Concurrently, the root quantity and length in transgenic NnWOX1-1 plants were greater than those in wild-type plants (Fig. 4a). In addition, transgenic NnWOX4-3 plants exhibited more roots, longer roots, and longer stems than wild-type plants (Fig. 4b). Although root length was greater in transgenic NnWOX5-1 plants than in wild-type plants, root quantity and stem height showed no significant differences (p < 0.05) between groups (Fig. 4c).

Functional analysis of NnWOX1-1, NnWOX4-3, and NnWOX5-1 in transgenic Arabidopsis plants. a Assessment of stem and root development in transgenic plants with constitutive NnWOX1-1 expression and wild-type plants. b Effect of NnWOX4-3 on stem growth and root development in transgenic Arabidopsis plants. c Effect of NnWOX5-1 on stem growth and root development in transgenic plants. The mean values were calculated from three replicate experiments, and error bars show standard deviation. Statistical analysis was performed using a Student's t-test, * P < 0.05
Transcriptome analysis of transgenic and wild-type plants
Transgenic Arabidopsis plants with NnWOX1-1, NnWOX4-3, and NnWOX5-1 and wild-type plants at the six-leaf stage were analyzed using RNA sequencing (RNA-seq) to monitor changes in gene expression. A total of 751, 594, and 541 upregulated genes (Additional file 1) were found in transgenic NnWOX1-1, NnWOX4-3, and NnWOX5-1 plants, respectively (Fig. 5a). These genes were classified into different biological pathways based on their functions. The most changes in gene expression occurred in general function prediction only, followed by signal transduction mechanism (Additional Fig. 3–1, 2, 3). The number of upregulated genes involved in plant hormone signal transduction pathways was the largest. There were 19, 19, and 13 upregulated genes involved in plant hormone signal transduction in the transgenic NnWOX1-1, NnWOX4-3, and NnWOX5-1 plants, respectively (Fig. 5b). All genes related to the ethylene and IAA signal transduction pathways and their corresponding factors were selected (Table 1). Two genes involved in ethylene synthesis (1-aminocyclopropane-1-carboxylate synthase and 1-aminocyclopropane-1-carboxylate oxidase homolog 8) were upregulated in all transgenic plants, which indicated that NnWOX1-1, NnWOX4-3, and NnWOX5-1 affected ethylene synthesis. The expression of some ethylene and IAA response factors (ethylene-responsive transcription factor ERF054, ethylene-responsive transcription factor ERF034, AP2/ERF and B3 domain-containing transcription repressor TEM1, ethylene-responsive transcription factor 14, AP2-like ethylene-responsive transcription factor SMZ, auxin-responsive protein IAA5, SAUR-like auxin-responsive protein, IAA-amino acid hydrolase ILR1-like 6, SAUR-like auxin-responsive protein, and IAA-amino acid hydrolase IAR3) was also enhanced in the transgenic NnWOX1-1, NnWOX4-3, and NnWOX5-1 plants (Table 1).

Statistical analysis of upregulated genes and pathway enrichment in transgenic plants with constitutive expression of NnWOX1-1, NnWOX4-3, and NnWOX5-1. a Number of upregulated genes following overexpression of NnWOX1-1, NnWOX4-3, and NnWOX5-1 in Arabidopsis plants. b Genes involved in plant hormone transduction pathway are counted in transgenic plants expressing NnWOX1-1, NnWOX4-3, and NnWOX5-1
Determination of ethylene, IAA, ABA, and lignin content
Ethylene, IAA, ABA, and lignin strongly influence AR formation in lotus plants; therefore, the levels of these compounds were measured in this experiment. Ethylene production rates were higher in NnWOX1-1, NnWOX4-3, and NnWOX5-1 transgenic plants than in wild-type plants (Fig. 6a), indicating that NnWOX1-1, NnWOX4-3, and NnWOX5-1 affected ethylene synthesis in transgenic plants. We also found that the IAA, ABA, and lignin contents in transgenic plants showed no significant differences compared to those in wild-type plants. The promotion of root development, stem growth, and flowering in transgenic plants expressing NnWOX1-1, NnWOX4-3, and NnWOX5-1 was probably related to ethylene synthesis (Fig. 6b–d).

Determination of ethylene, IAA, lignin, and ABA contents in transgenic plants expressing NnWOX1-1, NnWOX4-3, and NnWOX5-1 and in wild-type plants. a Ethylene content in six-leaf-stage transgenic and wild-type plants. b Analysis of IAA content in transgenic and wild type plants at the six-leaf stage. c Lignin contents of the three transgenic and wild-type plants were analyzed to monitor the effect of NnWOX1-1, NnWOX4-3, and NnWOX5-1 expression. d ABA content in NnWOX1-1, NnWOX4-3, and NnWOX5-1 transgenic and wild-type plants. For statistical analysis, the data were recorded as means ± SEs of three experiments with approximately 10 seedlings in each experiment. “*” indicates that values are significantly different between samples (P < 0.05)
Role of ethylene in AR formation and seedling growth in lotus plants
Lotus seedlings were treated with 300 mg/L of ethephon and 1-methylcyclopropene (1-MCP). The rate of AR emergence was significantly increased in the seedlings treated with ethephon and was dramatically inhibited following the application of 1-MCP. The rate of AR emergence under ethephon treatment reached an approximately maximum value (100%) at 6 d, whereas a rate of only 65% was observed in the 1-MCP treatment group (Fig. 7a). In addition, the stems of seedlings treated with ethephon were longer than those of the control and 1-MCP treatment group plants (Fig. 7b). Further experiments indicated that ethephon probably affected the accumulation of metabolites in lotus seedlings. Ethephon significantly promoted fresh and dry weights, and the highest fresh and dry weights were found in plants treated with ethephon for 25 d (Fig. 7c, d), which indicated greater availability of metabolites and energy during AR formation.

Role of ethylene in the development of lotus seedlings. a Effect of ethylene on the rates of adventitious root formation in lotus seedlings at 0, 2, 4, and 6 d after ethephon treatment. b Stem growth at 0, 2, 4, and 6 d after ethephon treatment in lotus seedlings. c Determination of fresh weight in response to ethephon treatment at 0, 5, 15, and 25 d in lotus seedlings. d Change in dry weight of lotus seedlings after ethephon treatment for 0, 5, 15, and 25 d. Each experiment was carried out with three replicates, and the data represent means ± SEs for approximately 20 seedlings. “*” indicates values that are significantly different between samples (P < 0.05)
Adaptation of transgenic Arabidopsis plants to drought and salt stress
Plants constitutively expressing NnWOX1-1, NnWOX4-3, and NnWOX5-1 were subjected to drought and salt stress to evaluate the functions of these three genes under adverse environmental conditions. Overexpression of NnWOX1-1, NnWOX4-3, and NnWOX5-1 improved the responses of transgenic plants to salt stress when compared with those of wild-type plants (Fig. 8a–c). In addition, NnWOX1-1 and NnWOX4-3 improved adaptation to drought resistance in the transgenic plants (Fig. 8a, b). Further analysis was performed to monitor gene expression in the three transgenic plant varieties. The expression of some stress-related genes (such as the MYB, WRKY, and N-acetylcysteine (NAC) transcription factors; and dehydration responsive element binding protein (DREB) and late embryogenesis abundant (LEA) proteins) was found to change in the transgenic plants when compared with that in the wild-type plants (Table 2). Therefore, the enhanced drought and salt stress tolerance observed in transgenic plants might be derived from the change in the expression of these genes.

Survival rates of transgenic NnWOX1-1, NnWOX4-3, and NnWOX5-1 plants and wild-type plants in response to drought and salt stress. a Survival rates of transgenic NnWOX1-1 Arabidopsis plants. b Survival rates of transgenic NnWOX4-3 plants and wild-type plants. c Survival rates of NnWOX5-1 transgenic plants. Data are presented as the means ± SE of three replicates, and p < 0.05 was accepted as the level of significance in statistical comparisons
Discussion
ARs are responsible for water and nutrient uptake and are necessary for plant growth, especially when principal roots are not thoroughly developed; thus, the developmental process of ARs affects the yield, quality, and adaptation of plants to the environment. Members of the WOX family are essential transcription factors that regulate various biological processes in plants. WOXs influence the apical meristem of roots, and the overexpression of these genes results in changes in root development [45]. For example, transgenic WOX11 and WOX12 plants exhibit more roots than non-transgenic plants [46, 47]. In a previous study, we screened transcripts from the entire genome of the lotus (Nelumbo nucifera Gaertn) and observed that the expression of some NnWOXs was induced by IAA and sucrose [23, 24]. In the present study, NnWOX1-1, NnWOX4-3, and NnWOX5-1 were cloned using RT-PCR (Fig. 1).
The WOX family is distributed among monocots and dicots, and several WOXs with superfamily homeodomains have been analyzed in detail within the past year. WOXs are grouped into three clades: ancient (distributed in algae and higher plants), intermediate (distributed in vascular plants), and modern (distributed in seed plants) [48, 49]. Among the 14 WOX proteins of Arabidopsis, WOX1-7 are classified into the modern clade; WOX8, 9, 11, and 12 belong to the intermediate clade; and WOX10, 13, and 14 belong to the ancient clade. The same distribution was also observed in an analysis of the gene structure of WOXs in poplars. According to the reports above, NnWOX1-1, NnWOX4-3, and NnWOX5-1 belong to the modern clade. Most WOXs in the modern clade have a homeodomain, a feature of the gene structure of this family in Arabidopsis [45]. In this study, we found that NnWOX1-1 and NnWOX4-3 contained a homeobox, whereas NnWOX5-1 contained a homeodomain. Therefore, NnWOX1-1 and NnWOX4-3 showed higher similarity to each other than to NnWOX5-1, which was further confirmed by the phylogenetic analysis of these three genes (Fig. 1). The homeodomain of Cucurbitaceae WOXs is highly conserved, suggesting that these genes have a close evolutionary history [50]. However, the lack of a homeodomain in NnWOX1-1 and NnWOX4-3 indicates that different functions might have existed in NnWOX1-1 and NnWOX4-3 than in NnWOX5-1. Members of the WOX family are diverse in gene structure, and the variation usually results in significant changes in biological processes. In our study, we found that 10 motifs were present in the three genes, with eight, five, and seven motifs included in NnWOX1-1, NnWOX4-3, and NnWOX5-1, respectively. This indicated that the functions of these three genes might be different in metabolic regulation mechanisms in lotus plants (Fig. 2).
Ethylene, a critical plant hormone, controls several metabolic processes, from seed germination to flowering. Ethylene positively regulates AR formation by controlling epidermal cell death before AR emergence [14]. In petunia cuttings, the development of ARs is regulated by ethylene [51]. The IAA signal is also strictly required for AR formation, as has been confirmed in lotus plants [23]. Sucrose also influences root development [52]. A further analysis of the effect of sucrose on AR formation showed that this effect mainly occurs during the induction period of AR development [53]. In lotus plants, the role of sucrose in AR formation has been assessed, and sucrose, like IAA, participates in AR development [25]. In the present study, the three genes were induced by IAA and sucrose, although their expression profiles were different. In addition, the upregulation of mRNA levels of NnWOX1-1, NnWOX4-3, and NnWOX5-1 varied in different organs (Fig. 3), suggesting that their functions in biological pathways might be different. This phenomenon is also found in Salix suchowensis; a total of 15 WOXs show organ-specific expression profiling, although some genes are classified into the same subgroup [54]. The transcriptional levels of WOXs in strawberry vary in roots, stems, and leaves, suggesting that these genes likely influence many developmental processes during plant growth by affecting gene expression [55].
The functions of NnWOX1-1, NnWOX4-3, and NnWOX5-1 were explored in Arabidopsis plants. We observed that transgenic plants expressing NnWOX1-1 and NnWOX4-3 had longer stems, more roots, and longer roots than wild-type plants, whereas the overexpression of NnWOX5-1 only improved root length (Fig. 4). WOX1 influences lateral organ formation [56], which has been further confirmed in Medicago truncatula [57]. A recent report stated that WOX1 can interact with auxins to regulate the development of leaf veins in cucumber plants [58]. Together with our findings, this suggests that WOX1 has multiple roles in plant growth. WOX4 is related to cytokinins and is required for meristem maintenance in rice [59]. In Arabidopsis, WOX4 influences the diverse functions of stem cells and procambium development [60]. PtWOX4 influences the auxin signal transduction pathway and regulates cell division in the stem [61]. In the present study, transgenic plants expressing NnWOX4-3 had enhanced root development and longer stems than wild-type plants (Fig. 4). We also found that the expression of many genes related to signal transduction mechanisms was changed in the three transgenic plants (Additional Fig. 3 - 1, 2, 3). Further analysis showed that genes involved in ethylene synthesis and response were upregulated (Table 1, Fig. 5). In addition, it was shown that the rates of ethylene synthesis were higher in the transgenic plants than in the wild-type plants (Fig. 6). This suggested that NnWOX1-1, NnWOX4-3, and NnWOX5-1, which promoted root development, were probably related to ethylene biosynthesis. Exogenous ethylene improves endogenous ethylene accumulation and auxin transport [21, 62], and auxin can also mediate ethylene to control AR formation in sunflowers [63]. Some important physiological indices showed that the lotus seedlings treated with ethephon in this study had increased AR emergence rates, stem heights, and fresh and dry weights (Fig. 7). Therefore, we concluded that ethylene signaling critically influences AR formation in lotus plants.
Responding to adverse environmental factors is very common for most members of the WOX family, suggesting that the expression of WOXs can improve plant adaptation to stress. Overexpression of OsWOX13 could improve drought tolerance in rice [64]. Several WOXs in cotton have been identified, and virus-induced gene silencing showed that these genes play a critical role in plant growth by responding to drought stress [65]. Han et al. reported that CsWOX, which is involved in the phytohormone pathway, plays multiple roles in plant growth and is responsive to stress [66]. The same phenomenon was found in our study, and constitutive expression of NnWOX1-1, NnWOX4-3, and NnWOX5-1 resulted in an improvement in salt tolerance in transgenic Arabidopsis plants (Fig. 8). However, further analysis was required to determine whether ethylene signaling affected drought and salt adaptation. Some stress-related genes, including DREB, MYB, WRKY, NAC domain containing protein, and LEA were found to have enhanced expression in the transgenic plants (Table 2). In previous studies, these genes have been shown to improve stress tolerance [67,68,69,70]. We believe that NnWOX1-1, NnWOX4-3, and NnWOX5-1 improve drought and salt tolerance in Arabidopsis plants probably by regulating expression of the abovementioned genes. Lignin synthesis strongly influences AR development via the sucrose pathway [26]. In this study, we found that the overexpression of NnWOX1-1, NnWOX4-3, and NnWOX5-1 did not affect lignin content in Arabidopsis plants, suggesting that the change in root growth did not result from the synthesis of IAA, ABA, and lignin. In summary, our findings provide primary insights into the roles of NnWOX1-1, NnWOX4-3, and NnWOX5-1. Further exploration of the regulatory network of AR formation and stress responses to lotus WOXs is necessary.
Conclusion
NnWOX1-1, NnWOX4-3, and NnWOX5-1 were cloned based on transcriptional information regarding AR development in lotus seedlings. The structures, phylogenetic relationships, motifs, and chromosomal locations were analyzed in detail. These three genes were induced by the exogenous application of IAA and sucrose. The constitutive expression of NnWOX1-1, NnWOX4-3, and NnWOX5-1 significantly promoted root development and stem growth in transgenic plants. Plant responses to stresses were also identified following transgene transfection in Arabidopsis. Further experiments confirmed that ethylene production rates in the transgenic plants were dramatically higher than those in wild-type plants. IAA, ABA, and lignin levels did not change, suggesting that NnWOX1-1, NnWOX4-3, and NnWOX5-1 regulated root formation in transgenic Arabidopsis plants probably through the biosynthesis of endogenous ethylene. We also found that ethylene had a positive effect on AR formation and growth in lotus seedlings. Our findings are helpful in further exploring WOX functions and providing a better understanding of AR formation in lotus plants.
Materials and methods
Preparation of plant materials
Lotus seeds (Taikong Lotus 36) were used to conduct experiments in this study. Seeds were obtained from The Guangchang Bailian Institute and cultivated in an experimental field of aquatic vegetables at Yangzhou University, Southeast China. For plant growth, the field was maintained at a 20–25 cm water depth in the spring, with a 40–50 cm water depth in the summer. The temperature was 25–35 °C during the day and 18–25 °C at night throughout the entire growing season (from April to October). Seeds were harvested in November and stored at room temperature.
Cloning of NnWOX1-1, NnWOX4-3, and NnWOX5-1
The full sequences of the three genes were obtained from the NCBI database. The following primers were used: NnWOX1-1, forward primer: 5′-ATGGAAAGGAAAGATGAT-3′, reverse primer: 5′-TCACTTGGATTTGAATGG-3′. NnWOX4-3, forward primer: 5′-ATGGAGGGCAAAGAGGAG-3′, reverse primer: 5′-TTACACTCTGGCCTTGAA-3′. NnWOX5-1, forward primer: 5′-ATGGGGAAGGACGTTGAG-3′, reverse primer 5′-TTAGACGTGAGGGTTGCT-3′. Total RNA was extracted using an RNeasy MinElute Cleanup Kit (QIAGEN, Germany), following the supplied instructions. The first cDNA strand was synthesized using 1–2 µg of total RNA following the elimination of DNA contamination. A 20 µL mixture containing 2.5 µL dNTP, 2 µL each of forward and reverse primers, 2.5 µL MgCl2, 2 µL Taq polymerase (5 U), 2 µL cDNA fragments, and 7 µL dH2O was utilized to perform PCR component analysis. PCR was performed using the following program: 1 min at 94 °C for initial denaturation; 35 cycles of 1 min at 94 °C for denaturation, 1 min at 56–60 °C for annealing, and 1 min at 72 °C for extension; and 10 min at 72 °C for the final extension. A GeneJET Gel Extraction Kit (Thermo, Germany) was used to extract target gene fragments, which were ligated into the cloning vector (pMD 18-T vector; TaKaRa). These ligated vectors were grown in DH5α Escherichia coli and sequenced by Sangon Biotechnology Co., Ltd. (Shanghai, China).
Sequence analysis
A comparison among these three genes was performed using the DNAman and Simple Modular Architecture Research Tool (SMART) software programs to analyze the conserved domains. NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins were selected, standardized to fasta format, and then successively imported into the software channels. In the option section, each sequence marked by black color was set to represent homology. A phylogenetic tree was constructed using DNAman and MEGA X software. In addition, the conserved motifs of the NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins were analyzed using the online DNAman and MEME server v5.4.1 software programs (http://meme-suite.org/tools/meme). First, standardized formats of NnWOX1-1-, NnWOX4-3-, and NnWOX5-1-encoded proteins were generated using DNAman software and then input into the data submission system of MEME software according to the instructions. Second, “zero-or-one occurrence per sequence (zoops)” and “ten” motifs in the required box were set in the option system. Finally, the output was visualized by Btools software. Chromosomal location maps of the three genes were constructed using DNAman and TBtools software. The detailed process has been described by Chen et al. [71].
Gene expression analysis
The RT-qPCR method was used to monitor the expression of the three genes in lotus seedlings treated with 20 g/L sucrose and 10 µL IAA. In addition, the ARs, leaves, stems of six-leaf seedlings, and flowers were selected to analyze organ-specific expression characteristics. Total plant RNA was extracted using an RNeasy MinElute Cleanup Kit (QIAGEN) according to the manufacturer's instructions. Genomic DNA was removed using DNase I, and then 3 µg RNA was used to synthesize first-strand cDNA with a RevertAid First Strand cDNA Synthesis Kit (Fermentas, USA). SuperReal PreMix Plus (Tiangen, China) was used to calculate the mRNA levels of NnWOX1-1, NnWOX4-3, and NnWOX5-1 with three biological replicates on an Mx 3000P machine (STRATAGENE, http://www.stratagene.com). The forward primer for NnWOX1-1 was 5′-GGTGGTGATTGTGAAGGAGT-3′, and the reverse primer was 5′-CAATCCAACGCCCTTACTAT-3′. The forward primer for NnWOX4-3 was 5′-TACTGTGGGTATTCAGGGCA-3′, and the reverse primer was 5′-TGACTCTTCTGGAAACCCTT-3′. The forward primer for NnWOX5-1 was 5′-CGGCTGTAACCTTTGGACTT-3′, and the reverse primer was 5′-TCCCAGGGCAGTTCCTTTTG-3′. Lotus β-actin was used as an internal standard, with the forward primer 5′-AACCTCCTCCTCATCGTACT-3′ and the reverse primer 5′-GACAGCATCAG CCATGTTCA-3′. The mRNA level was calculated using the 2−△△Ct method [21]. A 25 μL reaction mixture containing 12.5 μL SYBR Premix Ex Taq II (Tli RNaseH Plus) (2 ×); 1 μL of each primer (forward and reverse); 3 μL cDNA of NnWOX1-1, NnWOX4-3, and NnWOX5-1; and 8.5 μL dH2O was prepared. The PCR program comprised 30 s at 94 °C, followed by 40 cycles of 5 s at 95 °C and then 60 s at 60 °C.
Gene function analysis of NnWOX1-1, NnWOX4-3, and NnWOX5-1
NnWOX1-1, NnWOX4-3, and NnWOX5-1 were ligated into the pGEM-T vector (cloning vector) and sequenced by Sangon Biotechnology Co., Ltd., after being transferred into E. coli to amplify the plasmids. The three genes were removed from the cloning vector by digestion with BamHI and KpnI restriction enzymes and transferred into a plant transformation vector (pSN1301) containing the CaMV 35S promoter. After completing the vector construction process, pSN1301:: NnWOX1-1, pSN1301:: NnWOX4-3, and pSN1301:: NnWOX5-1 were transferred into Agrobacterium tumefaciens strain GV3101 for infection preparation. Wild-type Arabidopsis plants (Columbia) were transformed using the floral dip method [72]. The seeds of the T0 generation were screened using 20 μg/g hygromycin B on MS medium after sterilization. The screened plants were cultivated in a greenhouse at 22 °C under a 12 h photoperiod. Positive plants were further identified using RT-PCR. The primers, mixtures, and PCR processes for these three genes were the same as those used for gene cloning.
The T2 generation seeds of transgenic and wild-type plants at the six-leaf stage were selected for functional identification. First, the transgenic and wild-type seeds were sterilized with 70% alcohol for 10 s, and then sodium hypochlorite (10%) was added for 25 min. The seeds were placed on the medium and base material (soil:vermiculite, 1:1, v/v) for further analysis of plant phenotypes and root development.
RNA-seq analysis
RNA-seq of NnWOX1-1, NnWOX4-3, and NnWOX5-1
Transgenic and wild-type seeds were placed on the base material for germination and then transferred to a light house. The temperature was maintained at 22–23 °C with a 12 h photoperiod. The roots of six-leaf-stage plants were used for gene expression analysis by RNA-seq. Approximately 2–3 μg RNA of transgenic and wild-type plants was prepared for library construction. The detailed process for library construction is described by Cheng et al. [24], and sequencing was performed by Nan**g Jisi Huiyuan Biotechnology Co., Ltd., using a special construct.
Identification and annotation of differentially expressed genes (DEGs)
The samples were evaluated in parallel using Illumina gene expression sample preparation kits. DEGs were screened using the NOISeq method, and the detailed protocol was previously described by Cheng et al. [23]. The thresholds of the DEGs were fold change ≥ 2 and divergence probability ≥ 0.8. The genes obtained in this experiment were annotated using the Gene Ontology (GO) tool with three ontologies such as molecular function, cellular component, and biological process. All the DEGs were enriched and classified into various biological functions after comparison with the lotus genome obtained from the NCBI database. All the DEGs were compared to the GO database (http://www.geneontology.org/), and the number of genes was calculated for the three ontologies mentioned above. Further, these DEGs were then input into a list of significantly enriched GO terms determined by a hypergeometric test. For pathway (biological functions) analysis, the Kyoto Encyclopedia of Genes and Genomes (KEGG) tool was used for the organized enrichment analysis of DEGs. Specifically, all identified DEGs were grouped into different biological pathways following comparison with the genome data derived from the NCBI database using KEGG. Therefore, all the DEGs were grouped into different metabolic pathways.
Ethylene, IAA, and ABA identification
The transgenic NnWOX1-1, NnWOX4-3, NnWOX5-1, and wild-type seeds were cultivated on the base material and transferred into an illuminated incubator at 22 °C. Six-leaf plants were selected for ethylene, IAA, and ABA identification. First, three six-leaf-aged seedlings from each sample were washed and placed in a bottle tightly sealed with a plastic plug. These bottles were transferred to a dark carton for 10 d at 25 °C. The ethylene production rate was measured using a gas chromatograph (Agilent, USA) according to the procedure outlined by Fiserova et al. [73]. This experiment was repeated thrice. Then, 20 seedlings of the transgenic and wild-type plants were frozen in liquid nitrogen and ground into powder with a rod. Approximately 2 g of powdered sample from each plant was transferred into approximately 600 µL of isopropyl alcohol/water/concentrated hydrochloric acid (2:1:0.002 v/v/v) for IAA and ABA extraction. These mixtures were placed into an agitator for 25–30 min at 4 °C; 1 mL dichloromethane was added, and the shaking was repeated for 30 min at 4 °C. The mixtures were centrifuged at 10,000 rpm (5180 × g) for 5 min at 4 °C. The lower liquid layer was collected and dried under nitrogen. The dry powder was dissolved in 100 mL filtered methanol, and 50 mL of the resulting solution was subjected to liquid chromatography (Sigma, Germany) for IAA content determination [24]. ABA content was determined using the method reported by Zdunek and Lips [74].
Lignin identification
Thirty transgenic NnWOX1-1, NnWOX4-3, and NnWOX5-1 and wild-type plant roots were selected for polymer content analysis. The plant growth conditions were the same as those described in the “Gene function analysis of NnWOX1-1, NnWOX4-3, and NnWOX5-1” section. The plant roots were dried at 60 °C for approximately 2 d, and 60 mg dry power was transferred into ethanol (1 mL, 70%). The mixture was vortexed and centrifuged at 10,000 rpm at room temperature. The supernatant was discarded, and the precipitate was washed with 1 mL of acetone. A 1 mL solution of chloroform/methanol (1:1 v/v) was added to the precipitate after the supernatant was discarded. The precipitate was dried under vacuum. The precipitate was treated with 1.5 mL of 0.1 M sodium acetate buffer (pH 5.0) at 80 °C for approximately 30 min and then transferred into a tube with 10 µL of 0.01% sodium azide, amylase, and pullulanase and incubated at 37 °C overnight. After heating at 100 °C for a few minutes, the precipitate was washed with water multiple times. One milliliter of acetone was added to the sample, which was then dried via vacuum. The precipitate was dissolved in 100 µL of 25% acetyl bromide for 3 h at 50 °C. Four hundred microliters of 2 M sodium hydroxide and 70 µL of 0.5 M hydroxylamine hydrochloride were added. Acetic acid was added to increase the volume to 2 mL, and 200 µL of the supernatant was analyzed in a multifunctional microplate reader (1510–04201, Semefi, USA) to determine its absorbance at 280 nm. The polymer lignin content was calculated using the following formula:
(ABS, absorbance value; Coeff , absorption coefficient)
Stress responses for transgenic and wild-type plants
Approximately 50 seeds of transgenic and wild-type plants were placed in water for 24 h and then sown on the medium. The detailed cultivation conditions were the same as those described in “Materials and Methods section: Gene function analysis of NnWOX1-1, NnWOX4-3, and NnWOX5-1”. Six-leaf seedlings of wild-type and transgenic plants were subjected to drought and salt stress. For the drought treatment, the plants were first treated with a unified water management scheme, and then water was withdrawn for approximately 10 d. Survival rates were determined after 7 d of water recovery. For the salt treatment, 100 mM NaCl was used to treat transgenic and wild-type plants, and survival rates were determined after 10 d. For the survival rate, the maintenance of plant growth (some leaves remained green) or the occurrence of new leaves was used as a criterion for survival, and withering of plants during the recovery period was considered death. Three biological replicates were performed for each experiment, and at least 50 seedlings were used for each replicate.
Role of ethephon in AR formation and lotus plant growth
Approximately 300 lotus seeds were selected to identify AR formation rates. The seed coat was broken before treatment. Approximately 50 seedlings were first treated with 300 mg/L ethephon and 300 mg/L 1-MCP (1-MCP can bind ethylene receptors, preventing ethylene from affecting plant growth) for 2 d, and then transferred to water for continuous cultivation. For AR abundance analysis, 20 lotus seedlings cultivated for 0, 2, 4, and 6 d were selected to monitor AR formation. ARs with a length of≧0.2 cm after breaking through the hypocotyl were used to determine abundance. For stem length analysis, lotus seedlings cultivated for 0, 2, 4, and 6 d were selected to monitor growth status. A ruler graduated in centimeters was used to measure the stem length (from the hypocotyl to the bottom edge of the first leaf). For fresh and dry weight assessments, 20 seedlings cultivated for 0, 5, 15, and 25 d in water were used to monitor the change in growth. The surface water on the seedlings at each time point was first removed with absorbent material before weighing, and then the seedlings were dried in an oven at 60 °C. The dry weight was recorded after no change in the weight of the dried material was observed. Three replicates were carried out in the above experiments, including analysis of AR abundance, stem length, and fresh and dry weight determinations.
Statistical analysis
Statistical analyses were performed using Origin Pro 8.0 (Origin Inc., Massachusetts, USA). For the analysis in each experiment, three repeated experiments were carried out and the means ± SE of three repetitions were determined. The means with standard deviations are displayed in all figures. Significant differences were determined by Student's t-test and differences at p < 0.05 were accepted as the level of significance (*, p < 0.05).
Availability of data and materials
The material for all experiments was supported by the Aquatic Vegetable Lab of Yangzhou University. The collection of seeds complied with local and national guidelines and permissions were obtained. Detailed data have been deposited in the NCBI database (Project: PRJNA977099; CK1, CK2, CK3: SRR24775841, SRR24775840, SRR24775837; wox1-1, wox1-2, wox1-3: SRR24775836, SRR24775835, SRR24775834; wox4-1, wox4-2, wox4-3: SRR24775833, SRR24775832, SRR24775831; wox5-1, wox5-2, wox5-3: SRR24775830, SRR24775839, SRR24775838).
References
Shen-Miller J. Sacred lotus, the long-living fruits of China Antique. Seed Sci Res. 2002;14:131–43.
Ming R, VanBuren R, Liu YL, Yang M, Han YP, Li LT. Genome of the long-living sacred lotus (Nelumbo nucifera. Gaertn.). Genome Biol. 2013;14:R41.
Liu RX, Chen SM, Jiang JF, Zhu L, Zheng C, Han S, et al. Proteomic changes in the base of chrysanthemum cuttings during adventitious root formation. BMC Genom. 2013;14:919.
Borgi W, Ghedira K, Chouchane N. Antiinflammatory and analgesic activities of zizyphus lotus root barks. Fitoterapia. 2007;78:16–9.
Li SW, Leng Y, Feng L, Zeng XY. Involvement of abscisic acid in regulating antioxidative defense systems and IAA-oxidase activity and improving adventitious rooting in mung bean [Vigna radiata L. Wilczek] seedlings under cadmium stress. Environ Sci Pollut Res. 2014;21:525–37.
Falasca G, Zaghi D, Possenti M, Altamura MM. Adventitious root formation in Arabidopsis thaliana thin cell layers. Plant Cell Rep. 2004;23:17–25.
Kevers C, Hausman JF, Faivre-Rampant O, Evers D, Gaspar T. Hormonal control of adventitious rooting: progress and questions. J Appl Bot Angew Bot. 1997;71:71–9.
Li SW, Xue L, Xu S, Feng H, An L. Mediators, genes and signaling in adventitious rooting. Bot Rev. 2009; 75: 230–47.
Rasmussen A, Hosseini SA, Hajirezaei MR, Druege U, Geelen D. Adventitious rooting declines with the vegetative to reproductive switch and involves a changed auxin homeostasis. J Exp Bot. 2015;66:1437–52.
Teale WD, Paponov IA, Palme K. Auxin in action: signaling, transport and the control of plant growth and development. Nat Rev Mol Cell Biol. 2006;7:847–59.
E ZG, Ge L, Wang L. Molecular mechanism of adventitious root formation in rice. Plant Growth Regul. 2012;68:325–31.
Da Costa CT, De Almeida MR, Ruedell CM, Schwambach J, Maraschin FS, Fett-Neto AG. When stress and development go hand in hand: main hormonal controls of adventitious rooting in cuttings. Front Plant Sci. 2012;4:133.
Ahkami A, Scholz U, Steuernagel B, Strickert M, Haensch KT, Druege U, et al. Comprehensive transcriptome analysis unravels the existence of crucial genes regulating primary metabolism during adventitious root formation in petunia hybrida. PLoS One. 2014;9.
Mergemann H, Sauter M. Ethylene induces epidermal cell death at the site of adventitious root emergence in rice. Plant Physiol. 2000;124:609–14.
Rovere D, Fattorini L, Angeli SD, Veloccia A, Falasca G, Altamura MM. Auxin and cytokinin control formation of the quiescent centre in the adventitious root apex of Arabidopsis. Ann Bot. 2013;112:1395–407.
Mensuali-Sodi A, Panizza M, Tognoni F. Endogenous ethylene requirement for adventitious root induction and growth in tomato cotyledons and lavandinmicrocuttings in vitro. Plant Growth Regul. 1995;17:205–12.
Rovere FD, Fattorini L, Angeli SD, Veloccia A, Duca SD, Cai G, et al. Arabidopsis SHR and SCR transcription factors and AUX1 auxin influx carrier control the switch between adventitious rooting and xylogenesis in planta and in vitro cultured thin cell layers. Ann Bot. 2015;115:617–28.
Marchant A, Bhalerao R, Casimiro I, Eklöf J, Casero PJ, Bennett M, et al. AUX1 promotes lateral root formation by facilitating indole-3-acetic acid distribution between sink and source tissues in the Arabidopsis seedling. Plant Cell. 2002;14:589–97.
Friml J, Vieten A, Sauer M, Weijers D, Schwarz H, Hamann T, et al. Efflux-dependent auxin gradients establish the apical–basal axis of Arabidopsis. Nat. 2003;426:147–53.
Ljung K, Hull AK, Celenza J, Yamada M, Estelle M, Normanly J, et al. Sites and regulation of auxin biosynthesis in Arabidopsis roots. Plant Cell. 2005;17:1090–104.
Negi S, Sukumar P, Liu X, Cohen JD, Muday GK. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. Plant J. 2010;61:3–15.
Delarue M, Prinsen E, Onckelen VH, Caboche M, Bellini C. Sur2 mutations of Arabidopsis thaliana define a new locus involved in the control of auxin homeostasis. Plant J. 1998;14:603–11.
Cheng LB, Jiang RZ, Yang JJ, Xu XY, Zeng HT, Li SY. Transcriptome profiling reveals an IAA-regulated response to adventitious root formation in lotus seedling. Z. Naturforsch. 2018;73c;229–40.
Cheng LB, Han YY, Zhao MR, Li SY. Gene expression profiling reveals the effects of light on adventitious root formation in lotus seedlings (Nelumbo nucifera Gaertn.). BMC Genomics. 2020;21:707.
Cheng LB, Zhao MR, Hu ZB, Liu HY, Li SY. Comparative transcriptome analysis revealed the cooperative regulation of sucrose and IAA on adventitious root formation in lotus (Nelumbo nucifera Gaertn). BMC Genomics. 2020;21:653.
Cheng LB, Zhao C, Zhao MR, Han YY, Li SY. Lignin synthesis, affected by sucrose in lotus (Nelumbo nucifera) seedlings, was involved in regulation of root formation in the Arabidopsis thanliana. Int J Mol Sci. 2022;23:4.
Della Rovere F, Fattorini L, D’Angeli S, Veloccia A, Falasca G, Altamura MM. Auxin and cytokinin control formation of the quiescent centre in the adventitious root apex of Arabidopsis. Ann Bot. 2013;112:1395–407.
Sieberer T, Leyser O. Plant science-Auxin transport, but in which direction? Sci. 2006;312:858–60.
Xu M, Zhu L, Shou HX, Wu P. A PIN1 family gene, OsPIN1, involved in auxin-dependent adventitious root emergence and tillering in Rice. Plant Cell Physiol. 2005;46:1674–81.
Liu HJ, Wang SF, Yu XB, Yu J, He XW, Zhang SL, et al. ARL1, a LOB-domain protein required for adventitious root formation in rice. Plant J. 2005;43:47–56.
Park SO, Zheng Z, Oppenheimer DG, Hauser BA. The PRETTY FEW SEEDS2 gene encodes an Arabidopsis homeodomain protein that regulates ovule development. Development. 2005;132:841–9.
He P, Zhang YZ, Liu H, Yuan Y, Wang C, Yu JN, et al. Comprehensive analysis of WOX genes uncovers that WOX13 is involved in phytohormone-mediated fiber development in cotton. BMC Plant Biol. 2019;19:312.
Ji J, Strable J, Shimizu R, Koenig D, Sinha N, Scanlon MJ. WOX4 promotes procambial development. Plant Physiol. 2010;152:1346–56.
Haecker A, Gross-Hardt R, Geiges B, Sarkar A, Breuninger H, Herrmann M, et al. Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development. 2004;131:657–68.
Yang ZE, Gong Q, Qin WQ, Yang ZR, Cheng Y, Lu LL, et al. Genome-wide analysis of WOX genes in upland cotton and their expression pattern under different stresses. BMC Plant Biol. 2017;17:113.
Lopez-Moya F, Escudero N, Zavala-Gonzalez EA, Esteve-Bruna D, Blázquez MA, Alabadí D, et al. Induction of auxin biosynthesis and WOX5 repression mediate changes in root development in Arabidopsis exposed to chitosan. Sci Rep. 2017;7:16813.
Costanzo E, Trehin C, Vandenbussche M. The role of WOX genes in flower development. Ann Bot. 2014;114:1545–53.
Wang LQ, Wen SS, Wang R, Wang C, Gao B, Lu MZ. PagWOX11/12a activates PagCYP736A12 gene that facilitates salt tolerance in poplar. Plant Biotech J. 2021;19:2249–60.
Liu W, Xu L. Recruitment of IC-WOX genes in root evolution. Trends Plant Sci. 2018;23:490–6.
Nardmann J, Werr W. The invention of WUS-like stem cell-promoting functions in plants predates leptosporangiate ferns. Plant Mol Biol. 2012;78:123–34.
Yu J, Zhang YY, Liu W, Wang H, Wen ST, Zhang YJ, et al. Molecular evolution of auxin-mediated root initiation in plants. Mol Biol Evol. 2020;37:1387–93.
Sheng L, Hu X, Du Y, Zhang G, Huang H, Scheres B, et al. Non-canonical WOX11-mediated root branching contributes to plasticity in Arabidopsis root system architecture. Development. 2017;44:3126–33.
Li J, Zhang J, Jia H, Liu B, Sun P, Hu J, et al. The WUSCHEL-related homeobox 5a (PtoWOX5a) is involved in adventitious root development in poplar. Tree Physiol. 2018;38:139–53.
Liu BB, Wang L, Zhang J, Li JB, Zheng HQ, Chen J, et al. WUSCHEL-related Homeobox genes in Populus tomentosa: diversified expression patterns and a functional similarity in adventitious root formation. BMC Genomics. 2014;15:296.
Li XX, Hamyat M, Liu C, Ahmad S, Gao XM, Guo C, et al. Identification and characterization of the WOX family genes in five solanaceae species reveal their conserved roles in peptide signaling. Gene. 2018;9:260.
Xu M, **e W, Huang M. Two WUSCHEL-related HOMEOBOX genes, PeWOX11a and PeWOX11b, are involved in adventitious root formation of poplar. Physiol Plant. 2015;155:446–56.
Cheng S, Zhou DX, Zhao Y. WUSCHEL-related homeobox gene WOX11 increases rice drought resistance by controlling root hair formation and root system development. Plant Signal Behav. 2016;11: e1130198.
Zhang X, Zong J, Liu J, Yin J, Zhang D. Genome-wide analysis of WOX gene family in rice, sorghum, maize, Arabidopsis and poplar. J Integr Plant Biol. 2010;52:1016–26.
Zhou X, Guo Y, Zhao P, Sun MX. Comparative analysis of WUSCHEL-related homeobox genes revealed their parent-of-origin and cell type-specific expression pattern during early embryogenesis in tobacco. Front Plant Sci. 2018;9:311.
Zhang N, Huang X, Bao Y, Wang B, Liu LJ, Dai LJ, et al. Genome-wide identification and expression profiling of WUSCHEL-related homeobox (WOX) genes during adventitious shoot regeneration of watermelon (Citrullus lanatus). Acta Physiol Plant. 2015;37:224.
Druege U, Franken P, Lischewski SA, Ahkami AH, Zerche S, Hause B, et al. Transcriptomic analysis reveals ethylene as stimulator and auxin as regulator of adventitious root formation in petunia cuttings. Front Plant Sci. 2014;5:494.
Cano-Delgado AI, Metzlaff K, Bevan MW. The eli1 mutation reveals a link between cell expansion and secondary cell wall formation in Arabidopsis thaliana. Development. 2000;127:3395–405.
Takahashi F, Sato-Nara K, Kobayashi K, Suzuki M, Suzuki H. Sugar-induced adventitious roots in Arabidopsis seedlings. J Plant Res. 2003;116:83–91.
Wang XL, Bi CW, Wang CY, Ye QL, Yin T, Ye N. Genome-wide identification and characterization of WUSCHEL-related homeobox (WOX) genes in Salix suchowensis. J For Res. 2019;30:1811–22.
Cao YP, Han YH, Meng DD, Li GH, Li DH, Abdullah M, et al. Genome-wide analysis suggests the relaxed purifying selection affect the evolution of WOX genes in pyrus bretschneideri, prunus persica, prunus mume, and fragaria vesca. Front Genet. 2017;8:78.
Vandenbussche M, Horstman A, Zethof J, Koes R, Rijpkema AS, Gerats T. Differential recruitment of WOX transcription factors for lateral development and organ fusion in petunia and Arabidopsis. Plant Cell. 2009;21:2269–83.
Niu LF, Lin H, Zhang F, Watira TW, Li GF, Tang YH, et al. Loose flowering, a WUSCHEL-like Homeobox gene, is required for lateral fusion of floral organs in medicago truncatula. Plant J. 2015;81:480–92.
Wang H, Niu HH, Li C, Shen GY, Liu XF, Weng YQ, Wu T, Li Z. WUSCHEL-related homeobox1 (WOX1) regulates vein patterning and leaf size in Cucumis sativus. Hort Res. 2020;7:182.
Ohmori Y, Tanaka W, Kojim M, Sakakibara H, Hirano HY. WUSCHEL-RELATED HOMEOBOX4 is involved in meristem maintenance and is negatively regulated by the CLE gene FCP1 in rice. Plant Cell. 2013;25:229–41.
Ji JB, Shimizu R, Sinha N, Scanlon M. Analyses of WOX4 transgenics provide further evidence for the evolution of the WOX gene family during the regulation of diverse stem cell functions. Plant Signal Behav. 2014;5:916–20.
Kucukoglu M, Nilsson J, Zheng B, Chaabouni S, Nilsson O. WUSCHEL-RELATED HOMEOBOX4 (WOX4)-like genes regulate cambial cell division activity and secondary growth in Populus trees. New Phytol. 2017;215:642–57.
Cheng LB, Han YY, Liu HY, Jiang RZ, Li SY. Transcriptomic analysis reveals ethylene’s regulation involved in adventitious roots formation in lotus (Nelumbo nucifera. Gaertn). Acta Physiol Plant. 2019;41:97.
Liu JH, Reid DM. Auxin and ethylene-stimulated adventitious rooting in relation to tissue. J Exp Bot. 1992;43:1191–8.
Minh-Thu PT, Kim JS, Chae S, Jun KM, Lee GS, Kim DE. A WUSCHEL homeobox transcription factor, OsWOX13, enhances drought tolerance and triggers early flowering in rice. Mol Sci. 2018;41:781–98.
Tajo SM, Pan Z, He SP, Chen BJ, KM Y, Mahmood T, et al. Characterization of WOX genes revealed drought tolerance, callus induction, and tissue regeneration in Gossypium hirsutum. Front. Genet. 2022; 13:928055.
Han N, Tang R, Chen XG, Xu X, Ren ZH, Wang N. Genome-wide identification and characterization of WOX genes in Cucumis sativus. Genome. 2021;64:761–76.
Agarwal PK, Gupta K, Lopato S, Agarwal P. Dehydration responsive element binding transcription factors and their applications for the engineering of stress tolerance. J Exp Bot. 2017;68:2135–48.
Du P, Wang Q, Yuan DY, Chen SS, Su YN, Li L, Chen S et al. WRKY transcription factors and OBERON histone-binding proteins form complexes to balance plant growth and stress tolerance. EMBO J. 2023; e113639.
Lu M, Sun QP, Zhang DFb, Wang TY, Pan JB. Identification of 7 stress-related NAC transcription factor members in maize (Zea mays L.) and characterization of the expression pattern of these genes. Bioch BiophRes Commun. 2015; 462: 144e150.
Chourey K, Ramani S, Apte SK. Accumulation of LEA proteins in salt (NaCl) stressed young seedlings of rice (Oryza sativaL.) cultivar Bura Rata and their degradation during recovery from salinity stress. J Plant Physiol. 2003, 160; 1165–74.
Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, et al. Tbtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020;13:1194–202.
Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16:735–43.
Fiserova H, Mikusova Z, Klems M. Estimation of ethylene production and 1-aminocyclopropane-1-carboxylic acid content in plants by means of gas chromatography. Plant Soil Environ. 2008;54:55–60.
Zdunek E, Lips SH. Transport and accumulation rates of abscisic acid and aldehyde oxidase activity in Pisum sativum L. in response to suboptimal growth conditions. J. Exp. Bot. 2001; 52:1269–76.
Acknowledgements
We extend our thanks to members of Nan**g Jisi Huiyuan Biotechnology Co., Ltd. for their cooperation in obtaining the RNA-seq data during lotus AR formation. The authors also thank Editage for their editorial assistance.
Funding
This work was supported by the Cooperation Project of Yangzhou and Yangzhou University (YZ2020187) and the Modern Agricultural Development Project of Jiangsu Province (BE2021333). The funding bodies had no role in the design or performance of any experiments, data analysis, or writing of the manuscript.
Author information
Authors and Affiliations
Contributions
Cheng L. and Li S. conceived and designed the experiments. Liang S. and Han Y. performed the experiments. Zhao M. analyzed the data. Zhao C. and Liang S. analyzed the data. Cheng L. and Li S. wrote the text file. All authors have reviewed and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
We confirm that all the procedures were followed in accordance with the relevant national, international, and institutional guidelines. Consent to participate was not applicable to this study.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
quan, L., Shiting, L., Chen, Z. et al. NnWOX1-1, NnWOX4-3, and NnWOX5-1 of lotus (Nelumbo nucifera Gaertn)promote root formation and enhance stress tolerance in transgenic Arabidopsis thaliana. BMC Genomics 24, 719 (2023). https://doi.org/10.1186/s12864-023-09772-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-023-09772-w