
Abstract
Germline stem cell development and differentiation is tightly controlled by the surrounding somatic cells of the stem cell niche. In Drosophila females, cells of the niche emit various signals including Dpp and Wg to balance stem cell renewal and differentiation. Here, we show that the gene pzg is autonomously required in cells of the germline to sustain the interplay between niche and stem cells. Loss of pzg impairs stem cell differentiation and provokes the death of cells in the germarium. As a consequence of pzg loss, increased growth signalling activity predominantly of Dpp and Wg/Wnt, was observed, eventually disrupting the balance of germ cell self-renewal and differentiation. Whereas in the soma, apoptosis-induced compensatory growth is well established, the induction of self-renewal signals during oogenesis cannot compensate for dying germ cells, albeit inducing a new niche-like microenvironment. Instead, they impair the further development of germ cells and cause in addition a forward and feedback loop of cell death.
Similar content being viewed by others
Introduction
Stem cells maintain tissue homeostasis by a continuous and strictly controlled process of self-renewal and differentiation. In contrast to somatic cells, germline stem cells give rise to gametes with the ability to establish an entire organism upon fertilization, thereby passing genetic and epigenetic information from one generation to the next. Intricate developmental processes ensure the self-renewal and correct cellular differentiation of germline cells for a continuous supply of gametes and the protection of genome integrity. Drosophila oogenesis is a well-established model system to study those regulatory processes that are likely to apply widely to other organisms.
The adult Drosophila ovary consists of individual units named ovarioles, which harbour progressively developed eggs (for review1,2). At the anterior tip of each ovariole, two to three germline stem cells (GSCs) reside in a structure called the germarium, where they are directly associated with cells from somatic origin comprising the stem cell nicheFull size image
The close proximity to the centromere of the pzg locus at 78C5-C6 hampered the generation of FRT79B or FRT80B recombinants for further clonal analysis of pzg function in the germ line33. Instead, we specifically downregulated pzg activity during oogenesis with the help of small hairpin RNAs (shRNA) under UAS control using maternal Gal4-drivers32,34. Consistent with the data described above, two different shRNA-pzg constructs recapitulated the atrophied ovarian phenotype when induced with different germline specific Gal4-driver lines (Fig. 1c,c’; Fig. S1a–c). We concentrated on one each in the subsequent analyses, shRNA-pzg (BL35448) and nosGal4VP16 (BL4937). Efficient and germline-specific downregulation of pzg expression was verified in situ in adult ovaries, demonstrating that loss of nuclear Pzg protein follows the expression of the maternal Gal4-driver (Fig. 1e,f”). With this tool in hand, we further investigated the function of pzg in the female germline of Drosophila.
Depletion of maternal pzg activity affects germ cell differentiation
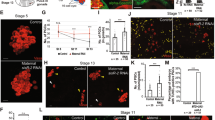
Depletion of pzg in germline cells generated ovaries containing only conical, germaria like structures, indicating that subsequent differentiation and development of egg chambers are hampered (Fig. 1c,c’). This may result from the absence of GSCs or from a failure of GSC maintenance and differentiation. GSCs can be identified by the expression of phosphorylated Mad (p-Smad), the activated form of Drosophila Smad, as well as by the GSC-specific organelle, the spherical spectrosome35,36,37. Immunostaining of ovaries from young females (0–3 days) for p-Smad reveals typically 2–3 GSCs in the control. In contrast, some of the shRNA-pzg depleted germaria contained only 0–1 GSCs within the niche. The majority, however, exhibited in addition a drastic increase in the number of p-Smad positive cells (Fig. 2a–c). Many of them were Vasa positive and contained spherical spectrosomes, typifying them as GSCs (Fig. 2b, see enlargements and Fig. S2). Many of the extra GSCs were no longer restricted to the anterior tip of the germarium, but were detected far posteriorly (Fig. 2b,c). In some cases, the apparent GSC clusters were surrounded by follicle cells giving a cyst-like appearance (Fig. 2b). Overall, loss of pzg apparently induced a type of a strongly restricted GSC tumour. The number of p-Smad positive cells decreased with age so that in 7 days old germaria barely any were detected (Fig. 2b).

Loss of pzg entails a surplus of undifferentiated GSCs. (a) In control germaria (nosGal4VP16::UAS-lacZ) of 0–3 days on average 2–3 GSCs can be detected by a strong p-Smad signal (green, arrows) and a spherical fusome (red with anti-Hts, appears pink in the overlay) only at the tip of the germarium. Cells of the germline are marked with anti-Vasa (V, blue). (b) The number of p-Smad positive cells is strongly elevated in 0–3 days old germaria where pzg activity was specifically depleted in cells of the germline (nosGal4VP16::UAS-shRNA-pzg). They are localized at next to the niche (closed arrows) and away from the niche (open arrows). Boxed area is magnified in the right panel: note that p-Smad positive cells (green, arrow) co-localize with Vasa (blue). Spectrosome can be discerned in some cells (asterisk; red, appears pink). In one week old germaria, the number of p-Smad positive cells is clearly decreased. (c) Quantification of GSCs per germarium in 0–3 days old nosGal4VP16::UAS-shRNA-pzg mutants (n = 25). Four categories were formed: no signal 0, 2–3, more (4–10) and >10 as indicated. The left column displays p-Smad positive cells within the niche. Only 24% of pzg depleted germaria display two to three p-Smad positive cells within the niche, a value typical for the control, whereas about 70% show more cells. The right column displays p-Smad positive cells outside of the niche. Note that more than half of the germaria show many more p-Smad positive cells mostly outside the niche. (d) bam-GFP expression (green; anti-GFP) was either absent (52%) or only marginally present (48%) in shRNA-pzg depleted germaria (anti-Vasa, blue, n = 25). Scale bars a-d: 25 μm, enlargement 5 μm.
Since ovaries depleted for Pzg do not develop maturing egg chambers, the GSCs may not carry on with the production of cystoblasts. In order to address the presence of cystoblasts and cystocysts, we studied the expression of bag of marbles (bam) using a bam-GFP reporter13. Depletion of Pzg in germline cells resulted in a near complete absence of bam-GFP signals. Only marginal and punctual signals were seen in about half of the germaria, whereas no signals were detected in the other half (Fig. 2d). This result is consistent with an elevated Dpp-signalling activity repressing bam expression, thereby impairing differentiation of germ cells lacking pzg. Apparently, pzg activity is required for a proper GSC cell lineage. Because the accumulation of GSCs was the most prominent defect, we focused on this phenotype in our further analyses.
Depletion of pzg activity in the germline increases Dpp-signalling activity
As the niche-derived Dpp-signalling activity plays a pivotal role in GSC self-renewal and proliferation, we sought for further evidence of a lost surveillance of BMP/Dpp-signalling activity in shRNA-pzg depleted germaria. In order to monitor dpp RNA expression levels by quantitative qRT-PCR in shRNA-pzg depleted ovaries, RNA was isolated from ovaries derived from freshly eclosed females to minimize the appearance of older egg chamber stages in the reference nosGal4VP16::UAS-lacZ. Nevertheless, control ovaries still occasionally harbour further developed egg chambers up to stage 6 (Supplemental Fig. S3a). To minimize effects of developmental stage differences between the shRNA-pzg depleted ovaries and control, we took great care in the selection of the reference genes dlp, Lamin C and slit: all three are expressed primarily in the somatic cells of the germarium, i.e. terminal filament cells, niche cells and FSCs4,38,39,40, and average mRNA levels were alike between pzg mutant and control (Supplemental Fig. S3b). Consistent with the higher number of p-Smad positive cells in situ, dpp mRNA levels were about 6-fold increased in shRNA-pzg depleted ovaries (Fig. 3a). Furthermore, expression of glass bottom boat (gbb) mRNA, encoding the second BMP-ligand of the stem cell niche6, was also elevated (Fig. 3a). We also assayed two targets of Dpp-signalling activity, bam and Dad, the former being repressed and the latter activated by Dpp6,41. Whereas expression of the Dpp target gene Dad was considerably enhanced, bam transcript levels were barely detectable (Fig. 3a), in agreement with the faint bam-GFP reporter activity in Pzg depleted ovaries (Fig. 2d). Overall, these results suggest that loss of pzg activity in germline cells provokes increased Dpp-signalling activity in the germarium, resulting in the retention of GSCs at the expense of the cystoblast daughter cell lineage.

Dpp-signalling activity is de-regulated in nosGal4VP16::UAS-shRNA-pzg mutant germaria. (a) qRT-PCR data gained from ovaries of freshly hatched females: In comparison to the control (nosGal4VP16::UAS-lacZ) the level of dpp, gbb and Dad transcripts is increased - whereas bam transcripts are strongly reduced in nosGal4VP16::UAS-shRNA-pzg depleted ovaries. Four biological and two technical replicates were performed. Three reference genes dlp, Lamin C and slit were used, and the amplification efficiencies taken into account for determining relative quantities by REST78. Median corresponds to expression ratio; mini-max depicts 95% confidence. Expression ratios shown were significant at the level of p < 0.05 using PFRR from REST: p = 0.0280 (dpp), p = 0.0595 (gbb), p = 0.0160 (Dad), p = 0.0200 (bam). (b) Rescue-assays aiming to deplete dpp activity either in the stem cell niche (using bab1-Gal4), in escort cells (with c587-Gal4) or in the GSC lineage (nosGal4VP16 and UAS-shRNA-dpp). The resulting ovaries of 3–5 days old females were subdivided into 5 categories according to their morphology after DAPI staining: 1: atrophied ovaries, only germaria like structures present; 2: atrophied ovaries with further developed stages up to stage 9; 3: development in some ovarioles proceeds up to stage 13; 4: fully developed eggs can be observed; 5: wild type ovaries. Scale bars represent 250 μm. Number of analyzed ovaries is given above each bar. Reduction of dpp activity in the niche or escort cells rescued the effect of shRNA-pzg induction in the germline lineage cells, to a lesser degree also in the germline derived lineage.
To directly test this hypothesis, we sought to downregulate dpp signals in shRNA-pzg mutant ovaries. Using specific Gal4-lines, dpp mRNA was depleted either in the somatic niche, the somatic escort cells, or within germline cells. Tissue specificity was ensured by using double-stranded RNAi of dpp under UAS control for somatic tissue (BL25782), and shRNA-dpp for the germline (BL36779). At least 100 ovaries per genotype were stained with DAPI to visualize their morphology and developmental progress within the ovarioles (Fig. 3b). To account for the range of phenotypes, we built five categories according to the developmental progress of egg chamber development. Downregulating dpp activity in the local microenvironment of the stem cell niche was achieved with the cap cell specific driver bric à brac (bab1)-Gal4, which on its own did not affect oogenesis. In contrast, dpp-RNAi induction in the cap cells of shRNA-pzg mutant ovaries resulted in a strong rescue of the atrophied phenotype: More than 50% of the ovaries contained ovarioles with further developed egg chambers, and in 20% even late egg stages developed (Fig. 3b). A similarly effective improvement was observed by using c587-Gal4, which reduced dpp levels within the surrounding escort cells6,36 (Fig. 3b). Germline specific depletion of dpp resulted in a much weaker phenotypic rescue, however, affected oogenesis itself to some degree (Fig. 3b). Together, these results indicate that pzg activity is required in the germline to restrict dpp levels in order to allow germ cell differentiation to proceed.
Pzg depletion affects GSC anchoring in the niche
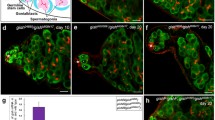
Our data suggest that loss of pzg increases Dpp-signalling activity, enforcing the continuous self-renewal of GSCs and impairing subsequent differentiation. Instead, GSCs agglomerate in the centre of the germarium as apparently undifferentiated GSCs18. In a wild type germarium, cell adhesion molecules like DE-Cadherin along the niche/GSC junction contribute to GSC self-renewal and differentiation, by anchoring the GSC within the niche and allowing asymmetric GSC-division to a new GSC and cystoblast daughter (for review42). Accordingly, Bam restricts the amount of DE-Cadherin in the differentiating cystoblast20. Earlier work has shown that differentiation-defective GSCs can outcompete healthy GSCs from the niche by occupying the limited contacts to the adjacent cap cells10. Altered niche contacts may therefore add an explanation for the observed defects in shRNA-pzg mutant germaria, like GSCs absence from the niche or ectopic GSCs away from the niche (Fig. 2b,c). In fact, the two cell adhesion molecules, DE-Cadherin and beta-catenin/Armadillo (Arm), showed an abnormal distribution. Whereas, Arm and DE-Cadherin concentrate at the junction between cap cells and GSCs in wild type germaria, both proteins accumulated to higher levels and often framed the p-Smad positive GSCs in shRNA-pzg mutant germaria (Fig. 4a,b’). Unbalanced levels of adhesion molecules may provoke a loss of GSCs from the niche proper, allowing their accumulation in the centre of the germarium42,43. In this case, we may expect to influence the shRNA-pzg mutant phenotype by modulating DE-Cadherin levels at the niche/stem cell junction. To this end, reduction of shotgun (shg), which encodes DE-Cadherin, was enforced by tissue specific RNAi. Downregulation of shg in the cap cells of the niche, but not in the escort cells, considerably improved the atrophied phenotype of shRNA-pzg mutant ovaries, allowing well matured egg chambers in about one third of the analyzed females, and even the development of late egg stages in a small fraction (Fig. 4c). Germline-specific downregulation of shg, however, had a very mild rescue effect with nearly half of the ovaries being completely atrophied (Fig. 4c). These results further support non-autonomous effects resulting from a loss of pzg activity within germline cells acting on somatic cells of the adjacent niche, thereby contributing to the inhibition of further GSC differentiation.

Level of cell adhesion molecules is increased and scattered in shRNA-pzg depleted germaria. (a,b’) Co-staining of p-Smad (green) and Arm (a,a’ magenta) or DE-Cadherin (b,b’ magenta) in 0–3 days old germaria using respective antibodies. In the control (a,b) nosGal4VP16::UAS-lacZ, Arm or DE-Cadherin is mainly detectable at the niche/GSC junction. In nosGal4VP16::UAS-shRNA-pzg germaria (a’,b’), Arm or DE-Cadherin is more enriched at the niche/GSC junction (arrows) and is additionally mislocalized encircling the GSCs (open arrows). Scale bars: 10 μm. (c) Rescue assays by changing levels of DE-Cadherin within the GSCs and non-autonomously in cells of the niche (using bab1-Gal4) or in escort cells (c587-Gal4) in otherwise pzg depleted germaria. Reducing the level of shg in the niche strongly ameliorates the shRNA-pzg atrophied phenotype, whereas concomitant reduction in escort cells (using c587-Gal4) did not. Reducing the level of shg within the GSC increases the number of atrophied ovaries of category 2. Genotypes of analyzed flies are given below. Categories of rescue are given in the legend. Number of analyzed ovaries is given above the bars.
Loss of pzg in germline cells provokes cell death in the germarium
Our results so far imply that a loss of pzg activity during germ cell development impairs differentiation by provoking an expanded Dpp-signalling activity and interfering with the adhesion properties of the niche/GSC junction. Both, the altered adhesion as well as increased Dpp activity, are reminiscent of answers from dying cells aiming to maintain tissue homeostasis (for overview43,44). Somatic cells undergoing apoptosis emit mitogenic signals, i.e. morphogens like Dpp, to stimulate compensatory proliferation in the surrounding tissue (for overview44). In fact, we already know that a loss of pzg during wing imaginal development not only induced apoptosis but also compensatory proliferation and apoptosis31. We therefore aimed to firmly establish that apoptosis of germline cells occurs as result of pzg depletion. We firstly applied Acridine Orange and TUNEL-staining as a hallmark of apoptosis. Indeed, even young (0–3 days) shRNA-pzg mutant ovaries strongly retain Acridine Orange dye (Fig. 5a,a”) and displayed several TUNEL-positive cells that were never observed in the control (Fig. 5b,b”). Some of these cells co-stained for p-Smad, whereas others were adjacent to p-Smad positive cells (Fig. 5b’,b”). In addition, the activated form of two different Drosophila effector Caspases, cleaved Caspase-3 and Dcp-1, were both observed in shRNA-pzg mutant germaria, whereas the control was devoid of specific Caspase activity (Fig. 5c,d”). Not only could we detect Caspase staining within or close to the niche within GSCs, but also in the cluster of undifferentiated GSCs located further posteriorly (Fig. 5c,d”). Caspase activity was not restricted to presumptive GSCs but was also detected in cells nearby (Fig. 5c’,d”). To determine whether pro-apoptotic gene activity of hid, reaper and sickle, one of the first indicators of cell death induction in Drosophila somatic cells, is likewise induced, we performed qRT-PCR analyses, which revealed an up to 5-fold induction or pro-apoptotic gene activity (Fig. 5i). In one week old germaria, signs of cell death and Caspase activity were still observed in shRNA-pzg mutant germaria, however not in the wild type control (Fig. 5e–h’). The number of dying germ cells, i.e. p-Smad positive cells or cells labelled by a spectrosome, declined, suggesting that they were finally eliminated (Fig. 5f–h’). Overexpression of the somatic Caspase inhibitors DIAP1 and/or p35 within the cells of the germline only slightly ameliorated the atrophied phenotype, allowing the development of a higher percentage of further developed egg stages up to stage 9 (Fig. S4). Unexpectedly, Acridine Orange and TUNEL staining as well as Dcp-1 Caspase activity were still observed in these germaria (Fig. S5), indicating that DIAP1/p35 overexpression cannot suppress cell death in the germ line. Either, the two may not be functional inhibitors of apoptosis and Caspase activity in the female germline and/or the observed cell death effects are not only based on apoptotic death but include other non-apoptotic mechanisms of cell death.

Loss of pzg triggers cell death in the germarium. Cell death was assayed in control (nosGal4VP16::UAS-lacZ; a–h) germaria and pzg-depleted germaria (nosGal4VP16::UAS-shRNA-pzg; a’–h’), of 0–3 day old females (a–d”) and of 7d old females (e–h’). (a,a”) Acridine Orange dye is selectively retained in dying cells; it is not detected in the control (a) but only in shRNA-pzg mutant germaria (a’,a”). (b,b”) TUNEL staining (magenta) marks DNA breaks within dying cells. No TUNEL signal can be detected in the control (b). TUNEL positive cells often co-localize with p-Smad positive cells (green) in the niche (encircled) and further posteriorly, but are also detected next to p-Smad positive cells in shRNA-pzg depleted germaria (b”, open arrow). Enlargements of boxed area are show to the right. (c-c”) Cleaved Caspase-3 and cleaved Dcp-1 activity (d-d”) (both in magenta) were observed only in pzg depleted germaria and not in the control. These signals were detected in GSCs labelled with Hts (green, encircled) next to the niche and in cell clusters more posteriorly, as well as in cells not labelled with Hts (open arrows). Co-staining with anti-Hts and Lamin C (green) was used to mark the spectrosomes/fusomes and the outlines of the cells). Enlargements of boxed area are shown to the right. Cell death is still detected in 7 days old germaria from shRNA-pzg depleted tissues (e’–h’), in contrast to the control (e–h). Cell death was detected with Acridine Orange (e-e’, green), TUNEL (f-f’, magenta), cleaved Caspase-3 (g-g’, magenta) and cleaved Dcp-1 (h-h’, magenta). GSCs were marked with p-Smad (f-f’, green), and with Hts (e-h’, green) marking the spectrosome (encircled). The majority of dying cells show no GSC specific marker (arrows). Enlargements of framed area are shown to the right. Scale bars: represent 25 μm, in enlargements 10 μm. (i) Quantitative RT-PCR revealed an induction of pro-apoptotic gene expression (reaper, hid and sickle) in shRNA-pzg mutant germaria. Four biological and two technical replicates were performed. Lamin C, dlp and slit served as reference genes. Median corresponds to expression ratio; mini-max depicts 95% confidence. All expression ratios shown were significant at the level of p < 0.05 using PFRR from REST: p = 0.0050 (rpr), p = 0.0240 (hid), p = 0.0010 (skl).
Loss of pzg in germline cells triggers the expression of growth promoting and regulating genes
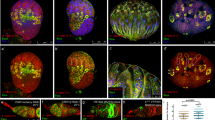
As shown above, shRNA-pzg mutant germ cells undergo cell death with typical signs of apoptosis. As a consequence, the dying germline cells may emit survival signals acting non-autonomously onto the surrounding cells. According to this working hypothesis, the observed rise in dpp and Dad expression (Fig. 3a), as well as the compromised adhesion properties (Fig. 4a,b’), might be a direct consequence of cell death induced by pzg depletion. In this case, we might expect the release of additional morphogens from the dying cells, as has been described for somatic tissue (for overview44,45). Indeed, immunostaining revealed a strong enrichment of Wg protein in cell groups that were in direct contact to the p-Smad positive cells, an effect never observed in the control (Fig. 6a). In accord with this, mRNA expression of wg and several other Wnt ligands known to promote proliferation in the germarium19,46 was elevated in pzg depleted ovaries as well (Fig. 6d). Likewise, we found significantly higher expression levels for hedgehog (hh) and Stat-lacZ (Fig. 6b,d). Moreover, RNAi-mediated depletion of Stat in either somatic niche cells or germline cells, occasionally allowed development of further developed egg chambers (Fig. S4). Inhibiting wg-signalling, particularly by overexpressing Axin (Axn) in escort cells, also improved the development of pzg depleted ovaries (Fig. S4). These rescue effects were not as strong as those observed by the reduction of dpp in the niche or in escort cells, emphasizing the central role of a deregulated Dpp-signalling as causative for the pzg-mediated ovarian atrophy.

Loss of pzg results in higher levels of growth regulatory factors. (a) Wg staining (red) is barely detectable in the control germaria (encircled; left panel); two GSCs (anti-pSmad, green, arrows) are located at the niche. In shRNA-pzg depleted germaria (right panels), Wg protein accumulates at higher levels and encircles the ectopic GSCs (stained with anti-pSmad, green, arrows). (b) Stat activity was detected with a Stat-lacZ reporter (anti-beta-Gal, red) in the cap cells of the niche (arrow) in control and pzg depleted germaria. Loss of pzg activity in the GSCs results in an increase in Stat-lacZ signals in the niche. Moreover, Stat-lacZ activity is detected also in cells surrounding the GSCs (open arrowheads. (c) In comparison to the control, puc-lacZ reporter activity (magenta, anti-beta-Gal) is increased in shRNA-pzg depleted germaria (arrows), notably framing p-Smad positive cells (green). Scale bars: 25 μm in all panels. (d) qRT-PCR revealed a considerable accumulation of wg, Wnt2, Wnt6, hh or eiger transcripts in nosGal4VP16::UAS-shRNA-pzg mutant germaria in comparison to the control (nosGal4VP16::UAS-lacZ). Four biological and two technical replicates were performed. Amplification efficiencies of the three reference genes dlp, Lamin C and slit were taken into account for determining relative quantities by REST. Median corresponds to expression ratio; mini-max depicts 95% confidence. All expression ratios shown were significant at the level of p < 0.05 using PFRR from REST: p = 0.0060 (wg), p = 0.0150 (Wnt2), p = 0.0190 (Wnt6), p = 0.0210 (hh), p = 0.0125 (eiger).
Overall, our results suggest that pzg depleted germline cells cannot differentiate but instead produce cell death-mediated mitogenic signals. Although these signals are important for the division and maintenance of GSCs in the female germline, we observed GSC loss with time. Perhaps the dying GSCs not only emit growth promoting but also other signals, which contribute to the final death of the cells in the germarium. Such a process is known from apoptotic somatic cells and relies on the production of the TNF orthologue Eiger by apoptotic cells47. Interestingly, we observed a significant induction of eiger expression by qRT-PCR in shRNA-pzg mutant ovaries (Fig. 6d). Moreover, an increase in JNK-signalling readout, visualized with puc-lacZ, was observed predominantly in cells surrounding p-Smad positive cells (Fig. 6c). As JNK-signalling is involved in various aspects of apoptosis including phagocytosis during oogenesis (for overview48), these results suggest induction of cell death in the germarium induced by Eiger and JNK-signalling activity as a consequence of pzg depletion. Together with the growth regulators induced as well, these two might contribute to the disturbed balance between proliferation- and differentiation in a pzg depleted germline (Fig. 7).

Model of pzg function in the germline. In a wild type ovary, short range Dpp-signalling activity is confined to the stem cell niche. Upon division, the daughter cell moves away from the niche thereby ensuring the upregulation of the differentiation gene bam. When pzg is depleted in germline cells, cellular homeostasis and stem cell niche communication is disturbed: As a consequence, cell death and growth promoting and regulating factors are induced. This provokes a new niche-like microenvironment inhibiting further differentiation of GSCs and finally the elimination of the undifferentiated, accumulated GSCs within the germarium.
