
Abstract
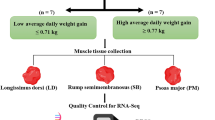
Sheep are considered one of the main sources of animal protein in Egypt and the producers of sheep mutton eagers to find biological criteria for selecting fast-growing lambs that reach market weight early. Therefore, the present study aimed to find a link between the expression profile of selected candidate genes with growth performance and carcass traits of Barki lambs. Thirty-eight Barki lambs were kept and fed individually after weaning till 12 months of age and were divided into 3 groups according to growth performance (fast, intermediate, and slow-growing). Three samples were taken from different body tissues (eye muscle, liver, and fat tail) of each group, directly during slaughtering and stored at − 80 °C until RNA isolation. Real-time PCR was used to profile selected candidate genes (RPL7, CTP1, FABP4, ADIPOQ, and CAPN3) and GAPDH was used as a housekee** gene. The results indicated that the final body weight was significantly (P ≤ 0.05) greater in the fast (49.9 kg) and intermediate (40.7 kg) compared to slow-growing animals (30.8 kg). The hot carcass weight was heavier (P ≤ 0.05) in the fast and intermediate-growing (24.57 and 19.07 kg) than slow-growing lambs (15.10 kg). The blood profiles of T3 and T4 hormones in addition to other parameters such as total protein, total lipids, and calcium level showed no clear variations among different experimental groups. At the molecular level, our data demonstrated upregulation of genes involved in protein biosynthesis (RPL7), fatty acid oxidation (CPT1), and lipolysis (FABP4) in the fast and intermediate-growing lambs in all studied tissues which facilitate protein accretion, energy expenditure, and fatty acid partitioning required for muscle building up. Moreover, the expression profile of the gene involved in muscle development (CAPN3) was increased in fast and intermediate-growing compared to slow-growing lambs in order to support muscle proper development. On the other hand, a candidate gene involved in lipogenesis (ADIPOQ) was expressed similarly in fat and liver tissues; however, its expression was increased in muscles of fast and intermediate-growing lambs compared to slow-growing animals. In conclusion, the current study indicated that the expression profile of genes involved in metabolic activities of liver, muscle, and adipose tissue is linked with the growth performance of lambs although no variations were detected in blood parameters. This provides an evidence for the importance of co-expression of these genes in body tissues to determine the final body weight and carcass characteristics of Barki sheep.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sheep are considered one of the main sources of animal protein. In addition, sheep can survive in the desert by grazing low-quality forage (Elshazly and Youngs 2019). Barki sheep are raised under a transhumant animal farming system in the Northwestern coastal desert of Egypt with a population of 470,000 heads (11% of the total Egyptian sheep population). In addition, this breed is well-known to be well-adapted to the harsh Egyptian desert conditions (El-Wakil and Elsayed, 2013). Generally, individuals of the same sheep breed are varied in growth performance and final body weight. Therefore, selecting lambs that grow fast and obtain heavy final body weight is crucial for improving the meat production industry, which is the main target of sheep breeding programs (Parker et al. 1991; Moghaddam et al. 2021).
Indeed, investigation of factors that control sheep growth based on monitoring live body weights over a relevant time is of major interest among scientists and meat producers (Lupi et al. 2015; Moghaddam et al. 2021). The growth performance of farm animals is an important economic trait that regulated by genetic and non-genetic factors (Alemneh and Getabalew 2019). In addition, the growth performance of sheep is influenced by type of birth, sex, breed, season, age, and pre-mating weight of the dam, which all are considered non-genetic factors (Yilmaz et al. 2007). Moreover, factors such as slaughtering animals at an early age (immature body weight), poor genetic potential for growth, and inappropriate feeding plan are major factors that reduce meat production (Kefelegn et al., 2019). Therefore, it is required to get all information on growth rate and the degree of live body weight maturity relevant to carcass composition for a genetic selection plan (Massender et al. 2019). For example, lamb birth weight is a determinant trait that influences the survivability and meat production performance of the sheep farm (Buzanskas et al. 2014; Ptáček et al. 2017).
Determination of genetic factors controlling phenotypic variation of productive traits in domestic sheep (Ovis aries) will facilitate efforts done for accelerating genetic improvement. Moreover, the identification of genes underlying sheep growth performance would support worldwide efforts in increasing mutton production (Wang et al. 2015). In this regard, gene expression profile was linked with the growth performance of Chinese (Miao et al. 2015) and Egyptian sheep breeds (Ashour et al. 2020; Miao et al. 2015). However, the available data that linked genetic variants and growth performance and carcass traits of Egyptian sheep is still limited. Therefore, investigating the transcriptional profile of muscle tissues would provide useful information to improve the production and quality of sheep meat (Chao et al. 2016; Sun et al. 2021).
Statistical analysis
Growth performance, carcass traits, biochemical measurements, and gene expression profile of selected candidate transcripts data were analyzed as a one-way analysis of variance using the SAS software, general linear model (v. 9.3, SAS Inst. Inc., Cary, NC, USA, 2011) as done by our group (Ghanem et al. 2014, 2021). The main effect was growth rate groups (fast, intermediate, and slow-growing animals). The following model was used: Yij = μ + Gi + eij, where:
-
Yij: The jth observation of the ith growth rate groups.
-
μ: The overall mean.
-
Gi: The effect of the ith growth rate groups (fast, intermediate, and slow-growing animals).
-
Eij: Random error.
All data are reported as least square means (LSM) ± standard errors (SE). Mean values were separated when significance existed; using Duncan’s multiple range test (Duncan’s, 1955). The significance level was set at 5%.
Results
Growth performance traits
The birth weight of the fast-growing animals was similar to that of slow and intermediate-growing lambs. The least square means of initial body weight, the final body weight, average daily gain, and total body gain are shown in Table 2 and Fig. 1. The initial and final body weights were heavier (P ≤ 0.05) in the fast-growing than intermediate and slow-growing lambs. In addition, the initial and final body weights were greater (P ≤ 0.05) in the intermediate than slow-growing animals as shown in Table 2 and Fig. 1. Subsequently, the average daily gain was increased (P ≤ 0.05) in the fast and intermediate-growing compared to slow-growing animals. Additionally, the total body gain was (P ≤ 0.05) greater in the fast-growing than slow-growing animals.

Growth rate of Barki lamb varied in growth performance under individual feeding management system
Carcass traits
The hot carcass weight (Table 3) was increased (P ≤ 0.05) in fast and intermediate compared to slow-growing lambs. Moreover, the liver weight of slaughtered lambs (Table 3) was significantly increased (P ≤ 0.05) in fast compared to intermediate and slow-growing lambs. On the other hand, dressing percentage, non-carcass fat, and tail fat were not significantly different among fast, intermediate, and slow-growing lambs. Furthermore, total body fat (non-carcass fat + tail fat kg) was increased but not significantly in fast and intermediate-growing compared to slow-growing lambs.
The blood profile of T3 and T4 hormone
The blood profile of thyroid hormones (T3 and T4) were not significantly (P ≤ 0.05) different among fast, intermediate, and slow-growing lambs (Table 4).
The blood profile of total protein
The total protein profile was not significantly different in fast, intermediate, and slow-growing lambs (Table 4).
The blood profile of total lipids
The level of total lipids was not significantly different in fast, intermediate, and slow-growing lambs (Table 4).
The blood profile of glucose level
The profile of blood glucose was not significantly different in fast, intermediate, and slow-growing lambs (Table 4).
The blood profile of calcium profile
The concentration of blood calcium was not significantly different in fast, intermediate, and slow-growing lambs (Table 4).
Gene expression profile of selected candidate transcripts
The expression profile of protein biosynthesis regulating candidate gene (RPL7) was significantly increased (P ≤ 0.05) in muscle (Longissmus dorsi) and liver tissues of fast and intermediate-growing compared to slow-growing lambs. On the other hand, adipose tissue of intermediate-growing animals recorded a higher expression profile of RPL7 gene than fast and slow-growing lambs (Fig. 2).

Expression profile of protein biosynthesis gene (RPL7) in different body tissues of Barki lambs varied in growth performance under individually feeding and management system
The transcript abundance of CPT1 which is involved in β-oxidation process was increased significantly in muscle and liver samples of fast-growing lambs than intermediate and slow-growing animals. In addition, the intermediate-growing lambs had greater transcript abundance than slow-growing group (Fig. 3). However, the expression of this gene in fat tissue was increased (P ≤ 0.05) in fast and intermediate-growing than slow-growing lambs.

Expression profile of lipolysis gene (CPT1) in different body tissues of Barki lambs varied in growth performance under individually feeding and management system
Noteworthy, transcript abundance of FABP4 (lipogenesis) was increased in all body tissues in fast and intermediate-growing compared with slow-growing lambs (Fig. 4).

Expression profile of lipogenesis gene (FABP4) in different body tissues of Barki lambs varied in growth performance under individually feeding and management system
The expression of ADIPOQ was increased (P ≤ 0.05) in muscle, liver, and fat tissues of fast and intermediate-growing compared to slow-growing lambs (Fig. 5). In addition, the adiponectin expression was higher in muscle collected from fast-growing than intermediate-growing lambs.

Expression profile of lipogenesis gene (ADIPOQ) in different body tissues of Barki lambs varied in growth performance under individually feeding and management system
The transcript abundance of CAPN3 was greater significantly in muscles of fast and intermediate than slow-growing animals (Fig. 6).

Expression profile CAPN3 in muscle of Barki lambs varied in growth performance under individually feeding and management system
Discussion
Growth traits such as birth weight, growth rate, body weight at slaughtering, and carcass characteristics of Barki sheep are crucial economic parameters. These parameters determine the productive efficiency of the animals to support the increased demand for mutton in Egypt (Sallam et al. 2012). Therefore, it is a privilege to select animals for meat production that grow fast and reached heavy bodies at slaughtering (Parker et al. 1991; Richards and Atkins 2004; Wang et al. 2015; Zhang et al. 2013a, b). The growth rate prior to weaning is considered a vital factor in determination of the production system performance. The fast-growing lambs, that quickly reach slaughter weight, will proportionately reduce the maintenance cost and increased farm revenue (Richards and Atkins 2004; Buzanskas et al. 2014; Wang et al. 2015; Zhang et al. 2013a, b).
Barki sheep is an Egyptian breed that survives well under the desert ecosystem, in addition to producing meat of high demand (El-Wakil and Elsayed 2013). The flock of Barki sheep of this investigation was under breeding program since 19,630 aiming to improve meat production (El-Wakil and Elsayed 2013). In the current study, the initial body weight of fast-growing lambs was greater than those intermediate and slow-growing lambs, which were positively linked with increased final body weight in fast-growing lambs. The total and average daily body gain were increased (P ≤ 0.05) in fast-growing lambs (22.3 kg and 173.6 gm) compared to intermediate (18.2 kg and 112.0 gm) and slow-growing lambs (15.0 kg and 92.3 gm). On the other hand, the growth rate of Awassi lambs was increased compared to Washera and Wollo highland breeds reflecting the genetic potentiality of body weight gain among these three breeds (Moghaddam et al. 2021).
Our results demonstrated that the final body weight of fast-growing animals was increased (49.9 kg) compared to intermediate (40.7 kg) and slow-growing groups (30.8 kg). The average body weights of Egyptian breeds were approximately 51 kg, 53 kg, and 44 kg at marketing age for Ossimi, Rahmani, and Barki breeds (Almahdy et al. 2000); Ashour et al. 2020; El-Malky et al. 2019; Hassan 2017; Khalifa et al. 2013; Marai et al. 2009; Miao et al. 2015). However, the body weights of Rahmani and Ossimi lambs were 40.8 and 36.6 kg at slaughtering under the same production system (Barkawi et al. 2009). In addition, daily weight gain and carcass traits of Awassi crossbred with Romanov were similar to that of pure Awassi breed (Tekel et al. 2020). On the other hand, Shaker et al. (2002) demonstrated that daily and total weight gain were increased in F1 lambs produced by crossbreeding Awassi ewes with Charollais and Romanov rams than the pure Awassi lambs. Indeed, individuals that grow fast with larger body size have greater economic returns; however, selection for lamb growth traits has been positively correlated with increased ewe mature body size (Borg et al., 2009; Posbergh and Huson 2021).
Birth weight of lambs is a key factor that influences growth performance of sheep (Buzanskas et al. 2014; Ptáček et al. 2017). The lamb final body weight is controlled by many genes such as MSTN, IGF-I, ADRB3, and leptin (Gholibeikifard et al, 2013). Within the muscle fibers, the biogenesis of ribosome and translation of protein are essential processes for the cells’ growth, differentiation, proliferation, and sha** animal development (Zhou et al. 2015). The building of muscle fibers and synthesis of protein required for animal growth are regulated partially by activity of ribosomal genes when proper nutrition is provided (Nader 2014). Indeed, the synthesis rate of muscle protein is mainly dependent on the content of ribosomes in muscle fibers (Millward et al. 1973). The expression profile of protein biosynthesis candidate gene (RPL7) was greater significantly in muscle (Longissmus dorsi) and liver tissues of fast and intermediate growing lambs than in slow-growing lambs which was in line with the increased body mass of fast and intermediate-growing lambs. Indeed, ribosomal proteins have a direct regulatory effect on the synthesis of protein that builds animal skeletal muscles (Costa et al. 2004; Han and Hickey 2005; Wang et al 2012). Protein is a crucial macronutrient participating in structural muscle building up in addition to its functional roles (hormones and enzymes) in the animal body (Mousa et al. 2019).
Moreover, it was found that the expression of RPL7 gene was downregulated in the broiler’s liver during feed restriction while there was no detection of differential expression of this gene in the adipose tissue (Wang et al. 2012). It was also reported that the genes involved in protein synthesis are altered severely due to diet restriction in porcine skeletal muscle (Costa et al. 2004; Han and Hickey 2005). Sheep genome-wide study proposed RPL7 as a candidate gene associated with growth performance and meat production traits (Zhang et al. 2013a, b). Additionally, RPL7 was linked with pre-weaning gain in German mutton merino (Wang et al. 2015). Noteworthy, our data indicated a higher expression profile of this gene in muscle and liver compared to adipose tissue. Thorrez et al. (2008) have demonstrated that the expression of RPL3 was enhanced in human adipose and muscle compared to liver cells. This spatiotemporal variation in gene expression between different species and tissues reflects changes of the cellular content of ribosomes and the activity of protein biosynthesis during different phases of body growth. In support of this idea, Cassar-Malek et al. (2007) indicated a change in the expression of ribosomal protein genes during different time points of bovine semitendinosus muscle development in the prenatal fetus stage.
Several studies have discovered genes associated with the post-weaning body weight of different sheep breeds (Gholizadeh et al. 2015; Al-Mamun et al. 2015). The upregulation of calpain3 (CAPN3) gene could be one of the main factors that enhanced the body mass of fast and intermediate-growing lambs more than the slow-growing animals. Calpains (CAPN) encode cysteine-activated intracellular proteases associated with increased number of myoblasts by controlling mitotic cycle (Barnoy et al. 1997; Cottin et al. 1994). Indeed, sheep individuals with TT genotype of CAPN had increased birth weight, final body weight, and average daily gain compared with other genotypes, while individuals with CC genotype recorded the lowest values of these traits (Mahrous et al. 2016). Several key transcripts involved in the development of skeletal muscles (MYL6B, MYH1, MYL3, MYO6, and MYOD1) were upregulated in the sheep fetal muscle that their maternal diet had high protein content (Sohel et al. 2020). This data indicates that increased dietary protein during pregnancy induces the expression of myogenic differentiation and development-related genes in the growing fetus.
The potential role of CAPN3 in enhancing sheep body weight has been reported by several authors who have demonstrated its genetic association with sheep growth performance and meat production traits (Koohmaraie 1992; Nassiry et al. 2007; Naveen et al. 2015). Therefore, it could be revealed that ribosomal and calpain encoded genes are co-expressed to enhance sheep body mass by stimulating myoblast proliferation and protein accumulation independent of the hormonal profile of thyroid hormones. Although increased plasma T4 concentration was recorded in Suffolk ewes compared to Gulf Coast native ewes, which was positively linked with larger body size and improved growth performance (Williams et al. 2004), however, our data did not show the same trend.
Animal body growth requires high-energy demand. In this regard, fatty acids are metabolized by an oxidation pathway in mitochondria to give acetyl-CoA, which is completely broken down via the Krebs cycle, or converted into acetate as an energy source (Guzman and Geelen 1993). Indeed, approximately 4–30% of circulating acetate and 10–55% of beta-hydroxybutyrate may be derived from hepatic oxidation of long-chain fatty acids in cattle (Mery et al. 1992). The catabolism of fatty acids occurs in the mitochondria by β-oxidation pathway (Sun et al. 2020). Our results demonstrated that the transcript abundance of CPT1 which is involved in the β-oxidation process was higher in muscle and liver samples of fast-growing lambs than intermediate and slow-growing lambs. Additionally, the intermediate-growing lambs had a greater transcript abundance of CPT1 than slow-growing lambs, while fast and intermediate-growing lambs had similar expression of this gene in fat tissue, although both had higher expression than slow-growing lambs.
A study that has been done in the fetal rabbit has shown increased expression of CPT I in cultured hepatocytes with long-chain fatty acids (Prip-Buus et al. 1995). The upregulation of this gene in fast-growing lambs may be due to increased metabolic demand to meet accelerated growth in fast and intermediate-growing lambs which is dependent on the utilization of fatty acids in both liver and develo** muscle fibers. A second candidate of mitochondrial β-oxidation genes is known as CPT1B which was downregulated in the skeletal muscle of offspring that their maternal dietary nutrition was restricted (Muroya et al. 2021). This may be due to reduced fatty acid metabolism as a result of decreased uptake of fatty acids. In support to the importance of β-oxidation genes, Price et al. (2003) reported a tenfold increase in the expression of mRNA encoding CPT1B in mammary gland cells of lactating compared to late pregnant and control ewes. In addition, downregulation of CPT1b reduced fatty acid metabolism and subsequently resulted in the obesity of rats (Warfel et al. 2017). Noteworthy, a putative SNP overlap** gene encoding mitochondrial gene known as ATP5F1A was related to progeny birth weight and litter means weight at birth of Iranian Baluchi sheep (Esmaeili-Fard et al. 2021).
Liu et al. (2021) detected a higher transcriptional profile of ADIPOQ and FABP4 in the subcutaneous fat (SCF) of Holstein than Korean Wagyu-cross steers, suggesting that there were more active adipocytes in the SCF of Holstein steers. Indeed, the expression of both FABP4 and ADIPOQ genes was upregulated during differentiation of adipocytes which indicated that adipocytes maturity in Wagyu-cross steers was more advanced than that in Holstein steers (Albrecht et al. 2011; Liu et al. 2021). Bionaz et al. (2012) indicated that Madin–Darby bovine kidney cells increased oxidation of long-chain fatty acid through upregulation of FABP4 gene in the cytoplasm and CPT1A in the mitochondria. Our data indicated a higher transcript abundance of FABP4 in all examined body tissues in fast and intermediate growing than slow-growing lambs. A genetic SNP in FABP4 gene was associated with meat tenderness in sheep (Xu et al. 2011). The data of this study strongly support the idea that oxidation of long-chain fatty acid is a key determining molecular mechanism explaining the variation of sheep growth performance. Additionally, the transcript abundance of ADIPOQ was higher in muscle, liver, and fat tissues of fast and intermediate growing than slow-growing lambs. Moreover, the adiponectin expression level in muscle collected from fast-growing was higher than intermediated growing lambs.
Adipokines have an important biological role in regulating lipid metabolism (Reynolds and Vickers 2019). The transcript abundance of ADIPOQ was higher in muscle, liver, and fat tissues of fast and intermediate growing than slow-growing lambs. In addition, the adiponectin expression level in muscle collected from fast-growing was higher than intermediated growing lambs. It was demonstrated that expression of ADIPOQ in fat collected 1 week after calving had a positive correlation with free fatty acid and subsequently the mobilization of adipose body reserves (Elis et al. 2013). In a recent study, a genetic association between the growth and carcass traits and the ADIPOQ haplotypes was detected on New Zealand Romney lambs (An et al. 2017). Similarly, the fast-growing and intermediate growing sheep had higher expression of the ADIPOQ gene which was linked with their growth performance. Moreover, a genomic variant (SNP) in ADIPOQ was associated with the marbling score of Hanwoo cattle (Shin and Chung 2013; Choi et al. 2015). Therefore, this study suggested that ADIPOQ gene could be used to differentiate lambs that varied in growth performance and carcass traits in addition to its role in regulating fat deposition and differentiation.
Conclusions
The data of the current investigation indicated that fast-growing lambs require upregulation of genes involved in protein biosynthesis (RPL7) which subsequently induced expression of muscle building up related transcript (CAPN3). The high demand of energy supply during muscle building up is provided by active transport of fatty acids through stimulation of FABP4 and ADIPOQ transcription. Finally, the utilization of fatty acids is performed inside mitochondria through β-oxidation process which is orchestrated by the CPT I gene. The co-expression of these genes in main body tissues is linked with growth performance and carcass traits of Barki lambs, which could be induced by the genetic makeup. However, this hypothesis requires a large population of lambs for validation.
Data availability
The data and materials of this study will be available on request.
Code availability
Not applicable.
References
Albrecht, E.; Gotoh, T.; Ebara, F.; Xu, J.X.; Viergutz, T.; Nürnberg, G.; Maak, S.; Wegner, J. 2011. Cellular conditions for intramuscular fat deposition in Japanese Black and Holstein steers. Meat Science, 89, 13–20.
Alemneh, T.; Getabalew, M. 2019. Factors Influencing the Growth and Development of Meat Animals. International Journal of Animal Science, 3, 1-5.
Almahdy, H.; Tess, M.W.; El-Tawil, E.; Shehata, E.; Mansour, H. 2000. Evaluation of Egyptian sheep production systems, II. Breeding objectives for purebred and composite breeds. Journal of Animal Science, 78, 288-295.
Al-Mamun, H.A.; Kwan, P.; Clark, S.A.; Ferdosi, M.H.; Tellam, R.; Gondro C.2015. Genome-wide association study of body weight in Australian Merino sheep reveals an orthologous region on OAR6 to human and bovine genomic regions affecting height and weight. Genetics Selection Evolution, 47, 66.
An, Q.; Zhou, H.; Hu, J.; Luo, Y.; Hickford, J.G.H. 2017. Haplotypes of the Ovine Adiponectin Gene and Their Association with Growth and Carcass Traits in New Zealand Romney Lambs. Genes (Basel), 8, 160.
Ashour, G.; Gad, A.; Fayed, A.K., Ashmawy, N.A.; El- Sayed, A. 2020. Evaluation of growth performance, blood metabolites and gene expression analysis in Egyptian sheep breeds, in relation to age. World Veterinary Journal, 10, 18-29.
Barkawi, A.H.; El-Asheeri, A.K.; Hafez, Y.M.; Ibrahim, M.A. Ali, M.M.. 2009. Growth and carcass characteristics of lambs in relation to plasma IGF-I and some histological traits of Longissimus lumbarum and Biceps femoris as affected by breed and age at slaughter. Livestock Science 124, 9–14.
Barnoy, S.; Glaser, T.; Kosower, N.S. 1997. Calpain and calpastatin in myoblast differentiation and fusion effects of inhibitors. Biochemical Biophysical Acta, 1358, 181-188.
Bionaz, M.; Thering, B.J.; Loor, J.J. 2012. Fine metabolic regulation in ruminants via nutrient-gene interactions, saturated long-chain fatty acids increase expression of genes involved in lipid metabolism and immune response partly through PPAR-α activation. British Journal of Nutrition, 107, 179-91.
Borg, R.C.; Notter, D.R.; Kott, R.W. 2009. Phenotypic and genetic associations between lamb growth traits and adult ewe body weights in western range sheep. Journal of Animal Science, 87, 3506–3514.
Buzanskas, M.E.; Grossi, D.A.; Ventura, R.V.; Schenkel, F.S.;Sargolzaei, M.; Meirelles, S.L.; Mokry, F.B.; Higa, R.H.; Mudadu, M.A.;daSilva, M.V.; Niciura, S.C.; Torres Jr, R.A.; Alencar, M.M.; Regitano, L.C.; Munari, D.P. 2014. Genome-wide association for growth traits in Canchim beef cattle. PLoS ONE, 9, 1-8
Cassar-Malek, I.; Passelaigue, F.; Bernard, C.; Leger, J.; Hocquette, J.F. 2007. Target genes of myostatin loss-of-function in muscles of late bovine fetuses. BMC Genomics, 8,63.
Chao, T.; Wang, G.; Wang, J.; Liu, Z.; Ji, Z.; Hou, L.; Zhang, C. 2016. Identification and classification of new transcripts in dorper and small-tailed han sheep skeletal muscle transcriptomes. PLoS One, 11,e0159638.
Choi, Y.; Davis, M.E.; Chung, H. 2015. Effects of genetic variants in the promoter region of the bovine adiponectin (ADIPOQ) gene on marbling of Hanwoo beef cattle. Meat Science, 105, 57-62.
Costa, N.D.; Mcgillivray, C.; Bai, Q.; Wood, J.D.; Evans, G.; Chang, K.C. 2004. Restriction of dietary energy and protein induces molecular changes in young porcine skeletal muscles. The journal of Nutrition, 134, 2191-2199.
Cottin, P.; Brutis, J.J.; Poussard, S.; Elamrani, N.; Broncard, S.; Ducastaing, A. 1994. Ca2_-dependent proteinases (calpains) and muscle cell differentiation. Biochimical Biophysical Acta, 1223, 170-178.
Duncan, D. B. (1955). "Multiple range and multiple F tests". Biometrics. 11 (1): 1–42. https://doi.org/10.2307/3001478. JSTOR 3001478
Elis, S.; Coyral-Castel, S.; Freret, S.; Cognié, J.; Desmarchais, A.; Fatet, A.; Rame, C.; Briant, E.; Maillard, V.; Dupont, J. 2013. Expression of adipokine and lipid metabolism genes in adipose tissue of dairy cows differing in a female fertility quantitative trait locus. Journal of Dairy Science; 9, 7591-602.
El-Malky, O.M.; Mostafa, T.H.; Ibrahim, N.H.; Younis, F.E.; Abd El-Salaam, A.M.; Tag El–Din, H.A. 2019. Comparison between productive and reproductive performance of Barki and Ossimi ewes under Egyptian conditions. Egyptian Journal of Sheep & Goat Sciences, 14, 61–82.
Elshazly, A.G.; Youngs, C.R. 2019. Feasibility of utilizing advanced reproductive technologies for sheep breeding in Egypt. Part 1. Genetic and nutritional resources. Egyptian Journal of Sheep and goat Sciences, 14, 39-52.
El-Wakil, S.; Elsayed, M. 2013. Genetic, phenotypic and environmental trends towards improving body weight in Barki sheep. Egyptian Journal of Sheep and goat Sciences, 8, 11-20.
Esmaeili-Fard, S.M.; Gholizadeh M.; Hafezian S.H.; and Abdollahi-Arpanahi, R. 2021. Genes and Pathways Affecting Sheep Productivity Traits, Genetic Parameters, Genome-Wide Association Map**, and Pathway Enrichment Analysis. Frontiers Genetics, 12,710613.
Frild, R.A.; Kemp, J.D.; and Varney, W.Y. 1961. Indices for lamb carcass composition. Journal of Animal Science, 322, 218.
Ghanem, N.; Ha, A.N.; Fakruzzaman, M.; Bang, J.I.; Lee, S.C.; Kong, I.K. 2014. Differential expression of selected candidate genes in bovine embryos produced in vitro and cultured with chemicals modulating lipid metabolism. Theriogenology, 82, 238-250.
Ghanem, N.; Ahmed, D.A.R.; Dessouki, S.M.; Faheem, M.S.; Gad, A.Y.; Peippo, J.; Barkawi, A.H. 2021. Cellular and molecular alterations of buffalo oocytes cultured under two different levels of oxygen tension during in vitro maturation. Zygote, 24, 1-11.
Ghasemi, M.; Zamani, P.; Vatankhah, M.; Abdoli, R. 2019. Genome-wide association study of birth weight in sheep. Animal, 13, 1797-1803.
Gholibeikifard, A.; Aminafshar, M.; Hosseinpour Mashhadi, M.. 2013. Polymorphism of IGF-I and ADRB3 genes and their association with growth traits in the Iranian Baluchi sheep. Journal of Agricultural Science and Technology, 15, 1153-1162.
Gholizadeh, M.; Rahimi-Mianji, G.; Nejati-Javaremi, A.; de Koning, D.J.; Jonas, E. 2015.Genome wide association study to detect QTL for body weight in Baluchi sheep. Journal of Genetics, 93, 489-493.
Guzman, M.; Geelen, M.J.H. 1993. Regulation of fatty acid oxidation in mammalian liver. Biochim. Biophys. Acta, 1167, 227-241.
Han, E.S.; Hickey, M. 2005. Microarray Evaluation of Dietary Restriction. The Journal of Nutrition, 135, 1343–1346.
Hassan, T.M.M. 2017. Effects of weaning age and some other factors on growth performance of Ossimi lambs. Egyptian Journal of Sheep & Goat Sciences, 12, 29,38.
Kefelegn, K.; Mekonnen, T.; Girma, A.; Goetsch AL. 2019. Analysis of Growth Performance Data in Sheep using Linear Mixed Model. World Jouenal of Agriculture and Soil Sciences, 2, 1-5.
Khalifa, A.I.; Ahmed, M.E.; Hafez, Y.H.; El-Zolaky, O.A.; Bahera, K.M.; Abido, A.A. 2013. Age at puberty and fertility of Rahmani sheep fed on biological inoculated corn silage. Annals of Agricultural Science, 58, 163–172.
Koohmaraie, M. 1992. Ovine skeletal muscle multi-catalytic proteinase complex (proteasome), purification, characterization, and comparison of its effects on myofibrils with mu-calpains. Journal of Animal Science, 70, 3697–3708.
Liu, L.; Cao, P.; Zhang, L.; Qi, M.; Wang, L.; Li, Z.; Shao, G.; Ding, L.; Zhao, X.; Zhao, X.; Xu, S.; Zhang, H.; Chai, J.; Yue, M.; Wang, G.; Liu, D.; Sun F. 2021. Comparisons of adipogenesis- and lipid metabolism-related gene expression levels in muscle, adipose tissue and liver from Wagyu-cross and Holstein steers. PLoS One,16, e0247559.
Lupi, T.M.; Nogales, S.; León, J.M.; Barba, C.; Delgado, J.V. 2015. Characterization of commercial and biological growth curves in the Segureña sheep breed. Animal, 9, 1341-1348.
Mahrous, K.F.; Hassanane, M.S.; Shafey, H.I.; Abdel Mordy, M.; Rushdi, H.E. 2016. Association between single nucleotide polymorphism in ovine Calpain gene and growth performance in three Egyptian sheep breeds. Journal of Genetic Engineering Biotechnology, 14, 233-240.
Marai, I.F.M.; Daader, A.H.; Bahgat, L.B. 2009. Performance traits of purebred Ossimi and Rahmani lambs and their crosses with Finnsheep born under two accelerated mating systems. Archives Animal Breeding, 52, 497-511.
Massender, E.; Brito, L.F.; Cánovas, A.; Baes, C.F.; Kennedy, D.; Schenkel, F.S. A. 2019. Genetic evaluation of growth, ultrasound, and carcass traits at alternative slaughter endpoints in crossbred heavy lambs. Journal of Animal Science, 97, 521-535.
Mery, R.S.; Liesman, J.S.; Herdt, T.H. 1992. Metabolism of long-chain fatty acids by ruminant liver. Journal of Nutrion, 122, 832-837.
Miao, X.; Luo, Q.; Qin, X. 2015. Genome-wide analysis reveals the differential regulations of mRNAs and miRNAs in Dorset and Small Tail Han sheep muscles. Gene, 562, 188–196.
Millward, D.J.; Garlick, P.J.; James, W.P.; Nnanyelugo, D.O.; Ryatt, J.S. 1973. Relationship between protein synthesis and RNA content in skeletal muscle. Nature, 241, 204-205.
Moghaddam, V.K.; Elahi, M.Y.; Nasri, M.H.F.; Elghandour, M.M.M.Y.; Monroy, J.C.; Salem, A.Z.M.; Karami, M.; Mlambo, V. 2021. Growth performance and carcass characteristics of finishing male lambs fed barberry pomace-containing diets. Animal Biotechnology, 32, 178-184.
Mousa, A.; Naqash, A.; Lim, S. 2019. Macronutrient and micronutrient intake during pregnancy, an overview of recent evidence. Nutrients, 11, 443.
Muroya S, Zhang Y, Kinoshita A, Otomaru K, Oshima K, Gotoh Y, Oshima I, Sano M, Roh S, Oe M, Ojima K, Gotoh T. 2021. Maternal Undernutrition during Pregnancy Alters Amino Acid Metabolism and Gene Expression Associated with Energy Metabolism and Angiogenesis in Fetal Calf Muscle. Metabolites, 11,582.
Nader, G.A.J. 2014. Ribosomes ‘muscle up’ postnatal muscle growth. Journal of Physiology, 592, 5143.
Nassiry, M.R.; Shahroudi, F.E.; Tahmoorespur, M.; Javadmanesh, A. 2007. Genetic variability and population structure in beta-lactoglobulin, calpastatin and calpain loci in Iranian Kurdi sheep. Pak. J. Biol. Sci, 10, 1062–1067.
Naveen, K.S.; Jayashankar, M.R.; Nagaraja, R.; Nagaraja, C.S.; Nadeem, F.; Satyanarayana, K. 2015. Genetic polymorphism of ovine Calpine gene in Bander sheep. International Journal of Scienticic Environmental Technology; 4, 804–812.
NRC. 1985. Nutrient requirements of sheep. 6th Edition, National Academy of Sciences, National Research Council, Washington, D.C.
Parker, W.J.; McCutcheon, S.N.; Wickham, G.A. 1991. Effect of administration and ruminal presence of chromic oxide controlled release capsules on herbage intake of sheep. New Zealand Journal of Agricultural Research (NZJAR), 34, 193-200.
Posbergh, C.J.; Huson, H.J. 2021. All sheeps and sizes, a genetic investigation of mature body size across sheep breeds reveals a polygenic nature. Animal Genetics 52, 99–107.
Price, N.T.; Jackson, V.N.; van der Leij, F.R.; Cameron, J.M.; Travers, M.T.; Bartelds, B.; Huijkman, N.C.; Zammit, V.A. 2003. Cloning and expression of the liver and muscle isoforms of ovine carnitine palmitoyltransferase 1, residues within the N-terminus of the muscle isoform influence the kinetic properties of the enzyme. Biochem Journal, 372, 871-879.
Prip-Buus, C.; Thumelin, S.; Chatelain, F.; Pegorier, J.P.; Girard, J. 1995. Hormonal and nutritional control of liver fatty acid oxidation and ketogenesis during development. Biochem. Soc. Trans, 23, 500-506.
Ptácek, M.;Duchácek, J.; Stádník, L.; Haki, J.; Fantová, M. 2017.Analysis of multivariate relations among birth weight, survivability traits, growth performance, and some important factors in Suffolk lambs. Archives Animal Breeding, 60, 43-50.
Reynolds, C.M.; Vickers, M.H. 2019. The role of adipokines in developmental programming, evidence from animal models. Journal of Endocrinology, 242, T81–T94.
Richards, J.S. ; Atkins, K.D., 2004. Simultaneous assortment of animals for meat and wool production in Merino flocks. Wool Technology and Sheep Breeding Journal, 52, 193–201.
Sallam, A.M.; Galal, S.; Rashed, M.A.; Alsheikh, S.M. 2012. Genetic diversity in Barki sheep breed in its native tract in Egypt. Egyptian. J. Anim. Prod., 49, 19–28.
SAS, Statistical analysis system institute. 2011. SAS institute, Inc., Cary, MC, USA.
Shaker, M.M. ; Abdullah, A.Y. ; Kridili, R.T. ; Balaha, J.; Sada, I.; Sovjak, R. 2002. Fattening performance and carcass value of Awassi, ram lambs, F1 crossbred of Romanov x Awassi and Charollais x Awassi in Jordan. Czech Journal of Animal Science, 47, 429-438.
Shin, S; Chung, E. 2013. Novel SNPs in the bovine ADIPOQ and PPARGC1A genes are associated with carcass traits in Hanwoo (Korean cattle). Molecular Biology Report, 40, 4651-60.
Sohel, M.M.H.; Akyuz, B.; Konca, Y.; Arslan, K.; Gurbulak, K.; Abay, M.; Kaliber, M.; Cinar, M.U. 2020. Differential protein input in the maternal diet alters the skeletal muscle transcriptome in fetal sheep. Mammalian Genome, 31, 309–324.
Sun, L.; Bai, M.; **ang, L.; Zhang, G.; Ma, W.; Jiang, H. 2016. Comparative transcriptome profiling of longissimus muscle tissues from Qianhua Mutton Merino and Small Tail Han sheep. Scientific Report, 6, 33586.
Sun, H.Z.; Zhou, M.; Wang, O.; Chen, Y.; Liu, J.X.; Guan, L.L. 2020. Multi-omics reveals functional genomic and metabolic mechanisms of milk production and quality in dairy cows. Bioinformatics, 36, 2530–2537.
Tekel1, N.; Baritci1, I.; Sireli1, H.D.; Tutkun1, M.; Eyduran, E.; Tariq, M. M. 2020. Determination of Fattening Performance and Carcass Characteristics of Awassi x (Romanov x Awassi) G1 Hybrid Male Lambs. Pakistan Journal of Zoology 52, 753-758.
Thorrez, L J.; Van Deun, K J.; Tranchevent, L.C J.; Van Lommel, L J.; Engelen, K J.; Marchal, K J.; Moreau, Y J.; Van Mechelen, I J.; Schuit, F. 2008. Using Ribosomal Protein Genes as Reference, A Tale of Caution. PLoS One, 3, e1854.
Wang, J.; Chen, W.; Kang, X.; Huang, Y.; Tian, Y.; Wang, Y. 2012. Identification of differentially expressed genes induced by energy restriction using annealing control primer system from the liver and adipose tissue of broilers. Poultry Science, 91, 972-978.
Wang, H.; Zhang, L.; Cao, J.; Wu, M.; Ma, X.; Liu, Z.; Liu, R.; Zhao, F.; Wei, C.; Du, L. 2015. Genome-Wide Specific Selection in Three Domestic sheep breeds. PLoS One, 10, e0128688.
Warfel, J.D.; Vandanmagsar, B.; Dubuisson, O.S.; Hodgeson, S.M.; Elks, C.M.; Ravussin, E.; Mynatt ,R.L. 2017. Examination of carnitine palmitoyltransferase 1 abundance in white adipose tissue, implications in obesity research. The American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 312, R816-R820.
Williams, C.C.; Calmes, K.J.; Fernandez, J.M.; Stanley, C.C.; Lovejoy, J.C.; Bateman, H.G.; Gentry, L.R.; Gantt, D.T.; Harding, G.D. 2004. Glucose metabolism and insulin sensitivity in Gulf Coast native and Suffolk ewes during late gestation and early lactation. Small Ruminant Research, 54, 167–171.
Xu, Q.L.; Tang G.W.; Zhang Q.L.; Huang Y.; KLiu., Y.X.; Quan K.; Zhu K.Y.; Zhang C.X. 2011. The FABP4 gene polymorphism is associated with meat tenderness in three Chinese native sheep breeds. Czech J. Anim. Sci., 56, 1–6.
Yilmaz, O.; Denk, H.; Bayram, D. 2007. Effects of lambing season, sex and birth type on growth performance in Norduz lambs. Small Ruminant Research, 68, 336–339.
Zhang, C.; Wang, G.; Wang, J.; Ji, Z.; Liu, Z.; Pi, X.; Chen, C. 2013a. Characterization and comparative analyses of muscle transcriptomes in Dorper and small-tailed Han sheep using RNA-Seq technique. PLoS One, 8, e72686.
Zhang, L.; Liu, J.; Zhao, F.; Ren, H.; Xu, L.; Lu, J.; Zhang, S.; Zhang, X.;, Wei, C.; Lu, G.; Zheng, Y.; Du, L. 2013b. Genome-wide association studies for growth and meat production traits in sheep. PLoS One, 8, e66569.
Zhang, C.; Wang, G.; Hou, L.; Ji, Z.; Wang, J. 2015. De novo assembly and characterization of the skeletal muscle transcriptome of sheep using Illumina paired-end sequencing. Biotechnology Letters , 37, 1747-56.
Zhou, X.; Liao, W.J.; Liao, J.M.; Liao, P.; Lu,H. 2015. Ribosomal proteins: functions beyond the ribosome. Journal of Molecular Cell Biology, 7, 92-104.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). The current study was partially supported financially by Desert Research Center, Cairo, Egypt.
Author information
Authors and Affiliations
Contributions
All authors of the present study have contributed equally by suggesting and designing the experiment in addition to writing and revising of the manuscript.
Corresponding author
Ethics declarations
Ethics approval
The Institutional Animal Care and Use Committee of Desert Research Center has accepted to conduct and approved this study.
Consent to participate
The authors of this work have agreed to participate in present study.
Consent for publication
The authors of this study have accepted to submit the current manuscript to Tropical Animal Health and Production.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ghanem, N., Zayed, M., Mohamed, I. et al. Co-expression of candidate genes regulating growth performance and carcass traits of Barki lambs in Egypt. Trop Anim Health Prod 54, 260 (2022). https://doi.org/10.1007/s11250-022-03263-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11250-022-03263-y