
Abstract
Context
Farmland biodiversity has been declining because of agricultural intensification and landscape simplification. Many farmland birds breeding in non-crop habitats use arable land as their feeding habitat (and vice versa) and understanding habitat composition and configuration at the landscape scale is important for their conservation.
Objectives
We explored the relationship between farmland bird densities and land-use characteristics at a landscape-scale (mean size 235 ha) to reveal the most important land-use elements driving avian farmland abundance.
Methods
We used bird territory map** from 36 study landscapes across Finland to study relationships between densities of total farmland birds, open field species, edge species, farmyard species, and Farmland Bird Indicator (FBI) species, and multiple descriptors of the composition and configuration of the study landscape mosaics, reflecting the full range of available crop types, farmland structures, non-crop habitat types, and soil type.
Results
Densities of farmland birds increased with greater areas of leys and pastures, subsidized grasslands, habitat diversity, and farmyards with animals, and those effects were consistently stronger compared to effects of non-crop habitats. Positive effects of the relative area of leys and pastures in the landscape was most often consistent in the species-specific models, whereas species-level responses to other landscape characteristics were idiosyncratic, reflecting the variety of the species’ ecologies and habitat requirements.
Conclusions
We demonstrate that overall habitat diversity, and habitat elements like subsidized grasslands, pastures, and farmsteads with animal production support higher bird diversity at the level of landscape mosaics. Our results suggest that studies based on field-scale study units need to be complemented with landscape-scale studies to reveal a holistic understanding of land-use intervention impacts on farmland birds.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Agricultural landscapes in Europe have undergone drastic simplification in the past century, mainly due to changing land-use practices aiming to maximise productivity (Robinson & Sutherland 2002). Consequently, natural and semi-natural habitats have been lost, resulting in overall landscape homogenisation (Benton et al. 2003; Maxwell et al. 2017; Tscharntke et al. 2012) with dramatic consequences for farmland biodiversity (Rigal et al. 2023). Population declines of farmland species have been particularly strong, which has been attributed to intensive farming practices (Donald et al. 2001; Rigal et al. 2023) and, more recently, climate change (Eglington & Pearce-Higgins 2012; Rigal et al. 2023). In general, a diverse landscape increases biodiversity, especially for birds, as many bird species depend on various habitat and field types to meet their needs in terms of foraging, nesting or wintering (Benton et al. 2003; Vickery & Arlettaz 2012). However, simplified crop rotations in arable systems have generally led to landscape homogenization (Newton 2017). Concomitantly with increasingly simplified crop rotation systems, the acreage of fallow lands has declined (Newton 2017) which is unfortunate since fallows and set asides can be the most beneficial agri-environment schemes (AES) for farmland species (Pe’er et al. 2017; Staggenborg & Anthes 2022).
Negative consequences of agricultural intensification on biodiversity motivated implementation of agri-environment schemes under the Common Agricultural Policy (CAP; Batáry et al. 2015), aiming to increase environmental sustainability in agricultural land-use. Yet in mosaic landscapes, both crop and non-crop elements contribute to landscape heterogeneity. Hedges, tree corridors, ditches and river verges, and in-field islets have been lost to facilitate modern farming with large machinery. Non-crop landscape structures are known to be of great value for farmland biodiversity (Bianchi et al. 2006), including bird populations (Batáry et al. 2010; Hiron et al. 2013a; Marja & Herzon 2012). Furthermore, farms with animal husbandry provide foraging grounds through manure heaps, paddocks and pastures (Hiron et al. 2013b; Šálek et al. 2020) and nesting sites in animal sheds, for example (Grüebler et al. 2010). However, the number of animal farms, especially smaller traditional ones, has drastically decreased over recent decades (-20% across the European Union; Eurostat 2023).
Farmland bird population trends have been recorded since the late 1970s throughout many European countries, including Finland, thanks to long-term, multi-national bird monitoring schemes (e.g., Brlík et al. 2021). Data availability, combined with the fact that birds serve as biodiversity indicators due to their spatially variegated habitat use and dependence on lower trophic levels (Gregory et al. 2005), led to the implementation of the Farmland Bird Indicator (FBI) in the 1990s. Today, the FBI is an integrated farmland biodiversity indicator in the Common Agricultural Policy and serves as one of the Structural and Sustainable Development Indicators, intended to provide a proxy for the wider biodiversity status in farmland habitats (Gregory et al. 2005; EEA 2013). Among the 39 species included in the European FBI, some species are more specialized in open habitats, or edge habitats such as hedges or forest edges, while others are dependent on farmland structures such as farmyards (Vepsäläinen et al. 2010). Thus, the FBI represents not only biodiversity related to open arable land but also to more complex and structured agricultural landscapes, which is especially pivotal in the context of landscape-scale analyses of farmland management effects on biodiversity. Since the establishment of the FBI, the indicator has decreased by 60% across Europe (PECBMS 2021a): forest birds, for example, showed a less dramatic decline of 10% (forest bird indicator; PECBMS 2021b).
Although effects of agricultural land-use and agri-environment schemes on farmland birds have been extensively studied, most studies have been conducted at a local scale (Kleijn et al. 2011; Siriwardena 2010). Relatively few studies have been conducted using a replicated landscape design, using entire landscapes as study units (Hiron et al. 2013a; Stjernman et al. 2019), even though landscape structure and land-use intensity are known to influence farmland biodiversity (Meier et al. 2022; Zingg et al. 2019). Because most farmland birds are central-place foragers during the breeding season, they typically rely on multiple habitat types to provide complementary and/or supplementary resources, such as food and nesting sites (Smith et al. 2014). Therefore, local-scale studies may miss important links between habitat types that ultimately matter for the breeding success and species-specific abundances of farmland birds at the landscape-scale – that is, in the interplay with other farmland types and features. Importantly, many farmland birds breeding in non-crop habitats, including nearby forests and farmsteads, use arable land as their primary feeding habitats (Tiainen & Pakkala 2001; Bruun & Smith 2003; Żmihorski et al. 2016), and understanding the effects of habitat composition and configuration at the landscape-scale is particularly important for such species. Mosaic agricultural landscapes, such as boreal agricultural regions, are ideal for studying such effects, because farmlands are typically concentrated into agricultural areas surrounded mainly by forests, making farmland landscapes physically isolated, independent entities.
In this study, we investigated the relationship between farmland bird densities and land-use characteristics at a landscape-scale in a northern European country, Finland. We focused on total densities of farmland birds; densities of birds breeding in open field habitats, edge habitats, and farmyard habitats; and abundances of individual species included in the Finnish FBI. Our research aimed to determine the relative importance of i) arable land-use and agri-environment scheme types, and ii) non-crop land-use, including non-crop habitats (e.g., in-field islets), farmland structures (e.g., farmyards), and landscape characteristics (such as soil type richness, habitat diversity and configuration) on landscape-level farmland bird assemblages. Broadly, we hypothesize the following responses, albeit such that not all three predictions will hold for all species or species guilds since they have different habitat requirements and specializations (e.g., open habitat species vs. edge species; summarized in Appendix Table S1).
-
i)
Species breeding in specific farmland habitats will benefit from larger areas of primary habitats (e.g., a larger area of open non-crop habitat and set asides will particularly benefit species breeding in open farmland, and farmyards will benefit farmyard and edge species).
-
ii)
Landscape complementation or supplementation across the landscape mosaic will affect farmland birds utilizing multiple habitats (e.g., an increasing area of set asides will benefit bird species breeding in edge or farmyard habitats but feeding in farmland; see Ekroos et al. 2019).
-
iii)
Increasing landscape heterogeneity will have positive effects on farmland bird assemblages and individual species for similar reasons as in ii), where heterogeneity is defined either directly by richness and diversity of land-use or indirectly via soil type richness (see, e.g., Vickery et al. 2009).
Materials & Methods
Study area
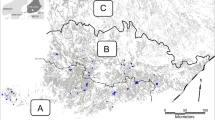
We analyzed effects of farmland management and the surrounding landscape characteristics on farmland birds at a landscape-level in 36 study landscapes located in South, Western and Central Finland (Fig. 1a). These 36 landscapes served as sample units for investigating bird densities and their associations with farmland habitats. These study areas were originally randomly selected, but the exact borders within which surveys were done were subsequently determined by the structure of the surrounding landscape. A natural feature of boreal farmland landscapes is that farmland occurs as patches within landscapes largely dominated by forests. Our study landscapes represent such farmland patches, rather than arbitrarily delineated landscape entities. Thus, the study areas were surrounded by continuous forest on most sides, which also determined the final size of each study area. The study landscapes also contained variable numbers and areas of non-farmland habitats (see Fig. 1). The average monitored area of the study landscapes was 235 ha (SD = 118 ha, range: 71–772 ha). We did not identify individual farms within the study landscapes, but the average farm size in Finland, based on utilized agricultural area, was 52 ha in 2022 (Official Statistics of Finland 2022). Field sizes varied from 0.42 to 2.22 ha. Fifteen study landscapes were visited in 2016–2017, while the remaining 21 landscapes were monitored in 2010 (see Ekroos et al. 2019).

Maps of the study area (a) with all study landscapes (grey points, N = 36) in South, West and Central Finland. Example study landscapes with low habitat diversity (b, upper red square in panel a), and with high habitat diversity (c, lower red square in panel a). Aggregated habitat classes are represented by distinct colors: arable and grasslands (beige/brown), farmland structures (blue), non-crop structures (yellow/green), landscape mosaic variables (green) and others (grey; see also Table 1). Note that the effective surveyed area covered all habitats except forests and clearcuts
Field surveys and data preparation
Bird territories within the study landscapes were monitored using the territory map** method, with three survey rounds from early May to mid-June. The territory map** was carried out by a team of experienced field ornithologists, where each team member surveyed slightly over 100 ha of farmland in one morning. All farmland habitats, including farmyards, settlements, and forest edges (but excluding the interior of forests and clearcuts) within the survey area were thoroughly searched for farmland birds, starting at sunrise, and ending roughly before noon. Territory observations were marked on visit maps, paying particular attention to documenting simultaneous observations of individuals from neighbouring territories. Based on the three-visit maps, we interpreted the positions of individual bird territories, which were subsequently represented as point objects in a GIS layer. In total, we found 54 bird species and recorded 11,414 territories (mean ± SD per landscape = 317 ± 167). One species (European golden plover Pluvialis apricaria) was omitted from further analyses due to its rare occurrence (N = 2). We first calculated the total number of territories per study landscape across all species, and thereafter the total bird density per hectare of surveyed farmland area (i.e., landscape area minus forest and clearcut area). We then classified bird species into different guilds based on their preferred habitats in Finland, following previous classifications (e.g., Tiainen & Pakkala 2001; Vepsäläinen et al. 2010): open field species (i.e., species that breed within open fields), edge species (i.e., species that breed in various non-crop habitats but frequently forage in farmland), and farmyard species (see SI Table S2 for species lists) and calculated their guild-specific densities per surveyed area. We also calculated species-specific abundances for the 14 species included in the Finnish FBI to investigate the species-specific patterns within the indicator (see Table 2).
To describe landscape characteristics, we used national digitized parcel maps (Integrated Administration and Control System database), supplemented with data on within-parcel boundaries of different crops based on aerial photographs and field map** during bird surveys. We also included GIS layers with digitized rivers, main drains, ditches, roads, forests, and islets (open, bushy or wooded), as well as farmyards and other built-up areas. These data were combined into one digitized vector map, providing spatially explicit, georeferenced data on the landscape characteristics and crop types (Fig. 1b, c).
We calculated multiple descriptors of the composition and structure of the study landscape mosaics, reflecting the full range of available crop types, farmland structures, non-crop habitat types, mean parcel size, and soil type (Table 1). We used summarized crop types instead of separating all crops individually, because broader crop type classes better reflect functional habitat types for farmland birds (Hiron et al. 2015).
Crop types included (i) leys and pastures (non-permanent, sown leys for silage and pastures on arable land that were grazed during the study year, often retained for some years as a part of crop rotation), (ii) fodder and hay (perennial and annual dry hay, silage and fresh fodder grasses, sown grasslands), (iii) spring-sown crops (cereals and dicots such as oilseed rapes, broad bean, etc.), (iv) autumn-sown crops (cereals, oilseed rape and caraway), (v) rotational set asides on arable land (set asides after crop**, i.e., stubble and open set aside), and (vi) subsidized grasslands (including a variety of environmental fallows, subsidized green manure crops, and seminatural grasslands or pastures), which together comprised all farmed land within each sampling unit. Farmyards and farmyards with animal husbandry (including dairy and beef cattle, horses, sheep, pigs, and poultry), individual barns not immediately connected to farmyards (historically used for fodder storage), and ditch verges were also mapped and digitized for inclusion in the analysis. We classified non-crop habitats into two classes, non-crop open (open and closed abandoned fields, and open and bushy in-field islets) and non-crop closed habitats (wooded islets, wooded ditch verges, and tree corridors).
Furthermore, to represent the overall land-use composition of the study landscapes, we also mapped clearcuts and forests, and calculated the relative area covered by farmland (total area of farmed land plus farmyards, barns, and set asides) per landscape. In addition, we calculated the habitat richness as the number of different habitat types per landscape, the habitat Shannon diversity index of all habitat types present in a landscape, and the farmland richness as the number of different farmland types per landscape (farmed land plus farmyards, barns, and set asides, see Table 1). These variables reflect the landscape's heterogeneity in terms of number of different (farmland) habitats present (i.e., richness) as well as the evenness of their proportional area in the study landscapes (i.e., Shannon diversity). Especially the latter can be seen as a proxy for landscape or resource complementation (Fahrig et al. 2011). We calculated the mean parcel size per landscape as the average area of each parcel across all habitat classes. For set asides, in-field islets, and leys and pastures, the relative number per landscape area was calculated for each and hereafter called patch density (PD; number of set asides or islets respectively divided by landscape area). To understand potential impacts of the soil composition, which may influence birds indirectly by varying fertility and vegetation structure due to soil properties, we used the freely available soil materials layer (GTK 2018) and extracted the soil type per parcel within each study landscape. Based on these data, we calculated the number of different soil types per study landscape.
In addition to the detailed habitat classes mapped directly in the field, we used CORINE landcover data (SYKE 2012) for information on the wider surroundings of each landscape. To do so we extracted the landcover data within a buffer zone of 5 km around the perimeter of each landscape. We were mainly interested in the area covered by forests and farmland types – which constitute the most abundant landcover classes besides waterbodies. Thus, we calculated the metrics percentage of landscape (PLAND in the R package landscapemetrics; Hesselbarth et al. 2019) and configuration (measured as mean distance between patches; ENN_MN in the same package) for forests (mixed, coniferous and deciduous forests aggregated from CORINE) and for the farmland classes arable land and pastures. See Table 1 for all predictors and Table 2 for all response variables used in the modelling.
Statistical analyses
All statistical analyses were performed in R (version 4.2.0, R Core Team 2022). For each response variable (total density, density per open field, edge and farmyard guild, density of FBI species, and FBI species-specific abundances) we followed the same statistical procedure. Predictors were log-transformed if necessary to account for non-linearity. In addition, the second order polynomial for area of farmyards, forests and ditches was tested and if near-significant (i.e., p < 0.1) included in the models to account for potential quadratic responses.
We first fitted single-predictor linear models for each explanatory variable described in Table 1. As the densities of the bird guilds had normally distributed residuals, simple linear models were fitted. For the species-specific models based on abundance, generalized linear mixed-effect models with a quasiPoisson error distribution were fitted and the log-transformed survey area per landscape was included as a fixed effect to account for varying survey areas. Predictors with p < 0.1 in single-predictor models were included in the full model. In case of collinearity (using a threshold of Pearson < 0.6) among predictors, we selected the one with better performance based on lower AIC values in single-predictor models. We avoided strongly correlated variables in the full models (Dormann et al. 2013), but we acknowledge that some landscape predictors are inherently correlated because of the nature of this observational study (SI Fig. S1-S2). Hence, we did not fit interactions between predictors. For the full model we ran a stepwise backwards model selection until only (near-)significant variables were left in the final model. We checked spatial autocorrelation for each model with none observed in the guild models. For the species-specific models, spatial autocorrelation was significant and thus glmmPQL models with a spatial error structure based on the landscape centroid coordinates were fitted for all 14 species (R package MASS, Venables & Ripley 2002). Model performance in terms of normality of residuals, heteroscedasticity, and outliers was checked for each final model (R package performance, Lüdecke et al. 2021). Specifically, outliers were checked based on a composite outlier score (‘check_outliers()’in the ‘performance’ R package, Lüdecke et al. 2021) obtained via the joint application of multiple outlier detection algorithms (median absolute deviation-based robust z scores, Leys et al. 2013; and Cook’s distance, Cook 1977). In case of detected outliers in either the explanatory or the response variables (for the species abundances), we reran the final model without the outliers (see SI Sects. 4.1 to 4.6 for details on outliers and sample sizes per model). In such cases, model estimates based on datasets without outliers are presented (Table 3, Fig. 2, Table 4).

Model-based linear regressions of most or second-most important predictor variable with a) total farmland bird density and habitat Shannon diversity index, b) open field species density and the percentage area of farmland, c) farmyard species density and the percentage area of farmyards with animals, d) edge species density and the area of farmland, e) FBI species density and the habitat type richness. Solid lines are the model estimates, shaded area the confidence intervals and points show the raw data distribution. Predictors of interest for the plots were extracted from the best model while other variables present were fixed at their means. The bar plots on the right of the regressions represent the relative variable importance per predictor from the best model per response variable
Results
On average, 1.73 ± 0.43 (mean ± SD) bird territories per ha for the surveyed farmland area were observed per landscape (Table 2). The overall density was 0.93 ± 0.29 territories per ha for the species included in the Finnish FBI. As divided into guilds, the highest densities were observed in edge species (0.53 ± 0.17), followed by open field species (0.51 ± 0.20), and farmyard species (0.28 ± 0.14). The three most abundant FBI species were skylark (Alauda arvensis; 55.39 ± 58.42 territories per study landscape), fieldfare (Turdus pilaris; 34.83 ± 30.87), and common whitethroat (Sylvia communis; 16.06 ± 11.02). The three rarest FBI species were corncrake (Crex crex; 0.81 ± 1.21), ortolan bunting (Emberiza hortulana; 3.03 ± 5.56), and jackdaw (Corvus monedula; 3.50 ± 4.28). See Table 2 for detailed densities per guild and species.
Community level responses
The overall density of all observed birds per ha for the farmland area showed significant positive relationships with certain landscape features and landscape configuration metrics. A larger mean distance between forest patches in the wider landscape (5 km buffer area) was negatively associated with the total bird density (model predicted estimate ± SE: -0.170 ± 0.057, p = 0.006), while the habitat Shannon diversity (at the scale of study landscapes) showed a strong positive effect (0.222 ± 0.063, p = 0.001; Fig. 2a). Greater areas covered by farmland (log-transformed estimates: -1.983 ± 0.913, p = 0.038) were linked to decreasing total bird densities, while a greater area of spring-sown crops showed a positive correlation (0.173 ± 0.063, p = 0.01; Table 3; SI Fig. S3).
Regarding open field species, a greater area of farmland was highly positively significant (0.102 ± 0.024, p < 0.001), along with greater area of pastures in the wider surroundings (5 km; 0.095 ± 0.025, p < 0.001; Table 3; Fig. 2b; SI Fig. S4). The area of fodder and hay fields was included in the best model, but the effect was non-significant (SI Table S3).
The density of farmyard species increased significantly with greater area of farmyards with animal husbandry (log-transformed estimates: 0.396 ± 0.110, p = 0.001), farmyards (log-transformed estimates: 0.234 ± 0.078, p = 0.005), and leys and pastures (log-transformed estimates: 0.124 ± 0.049, p = 0.047, p = 0.017; Table 3; Fig. 2c; SI Fig. S5).
Edge species’ density significantly decreased with larger areas of farmland (-0.096 ± 0.023, p < 0.001), while it increased with greater mean distance among pastures in the wider 5 km surroundings (log-transformed estimates: 0.139 ± 0.039, p = 0.002), and greater patch density of set asides within the study landscapes (log-transformed estimates: 0.279 ± 0.121, p = 0.028; Table 3, Fig. 2d; SI Fig. S6). The distance between forest patches (5 km) was included in the best model but had only a near-significant relationship (SI Table S3).
FBI species’ densities were significantly and positively related to habitat richness (log-transformed estimates: 2.299 ± 0.937, p = 0.020; Fig. 2e; SI Fig. S7), subsidized grasslands (log-transformed estimates: 0.352 ± 0.109, p = 0.003), and leys and pastures (log-transformed estimates: 0.245 ± 0.113, p = 0.039), while ditches were negatively associated (-0.100 ± 0.038, p = 0.014; Table 3). The variance explained (adjusted R2) of best models was generally high, ranging from 49 to 59% (Table 3).
Species-specific responses
The responses of individual Finnish FBI species were rather heterogeneous and sometimes contrasting (Table 4). Several landscape features were significant predictors of species-specific bird abundances. First, greater relative area covered by leys and pastures positively influenced the abundance of the lapwing (Vanellus vanellus), fieldfare, barn swallow (Hirundo rustica), and tree sparrow (Passer montanus; Table 4; SI Fig. S15d, S18c, S14a, S17c), while subsidized grasslands were positively linked with meadow pipit (Anthus pratensis; SI Fig. S9a) and tree sparrow (SI Fig. S17a) abundances. In addition, among variables describing individual farmland habitats, greater area of direct-sown spring crops showed a positive relationship with skylark abundance (SI Fig. S8b), whereas greater areas of spring-sown crops was negative for corncrake abundance (SI Fig. S19). Larger area of fodder and hay fields were negatively associated with skylark abundances (SI Fig. S8c).
Second, a greater relative area of farmyards with animals in the study landscapes was significantly associated with a higher abundance of the tree sparrow (SI Fig. S17b), and had a negative curvilinear link with the house martin (Delichon urbicum; linear term positive, quadratic term negative; SI Fig. S10), while larger areas of farmyards was positively related to fieldfare abundance (SI Fig. S18b) but negatively with lapwing abundance (SI Fig. S15c). Larger relative area of open non-crop habitats was significantly and positively related to the abundance of the fieldfare (SI Fig. S18a) whereas greater areas of closed non-crop habitats had negative effects on meadow pipits (SI Fig. S9b). Furthermore, amongst non-crop habitats, in-field islets were positively influencing the number of whitethroats (SI Fig. S13a), a larger area of barns was associated with higher tree sparrow abundance (SI Fig. S17d), and ditches positively with whinchats (Saxicola rubetra; SI Fig. S12b).
Lastly, several variables describing the landscape mosaic influenced individual FBI species. Habitat richness and diversity significantly influenced the abundances of three species: a negative association between habitat richness and lapwing abundance, a positive association between richness of farmland types and barn swallow abundance, and a positive association between habitat Shannon diversity and jackdaw abundance (SI Figs. S15e, S14b, and S21a). A larger relative area of clearcuts within the study landscapes was negatively associated with skylark and whitethroat abundances (SI Fig. S13a, S16b). Larger distances between forest patches within a 5 km buffer area were linked to lower abundances of whinchats and whitethroats (Table 3; SI Fig. S12a and S13c), while the relative forest area within 5 km was negatively linked to lapwings (SI Fig. S15a) and curlews (Numenius arquata; SI Fig. S16b), and showed a curvilinear link with jackdaws (linear term positive, quadratic term negative; SI Fig. S21b). The relative arable area in the wider surroundings was the only (negative) predictor of starling abundance (Sturnus vulgaris; SI Fig. S20), whereas a shorter mean distance between arable patches within 5 km was positively related to jackdaw (SI Fig. S21c) and curlew abundances (SI Fig. S16a). In contrast, shorter mean distances between pastures was positively associated with lapwing abundance (SI Fig. S15b). Finally, a larger relative farmland area was positively correlated with ortolan bunting abundance (SI Fig. S11), but negatively with fieldfare abundance (SI Fig. S18d). Explained variances (marginal R2) showed a wide range, from 14.4% (corncrake) to 80.7% (tree sparrow), with lowest values for corncrakes, whitethroats, starlings, and whinchats.
Discussion
Utilizing a large-scale study system spanning over 500 km latitudinally, and using entire landscapes as study units, we demonstrate landscape-level effects of local-scale agricultural land-use on farmland bird assemblages. Many of these effects are challenging to capture when using smaller sampling units, designed to provide direct comparisons between local-scale interventions and control areas, because many farmland birds use larger areas than individual fields for breeding and foraging. Thus, our results can be interpreted either through effects of larger areas of individual land-use types or interventions per se, or as combined landscape-wide effects through landscape supplementation or complementation. Understanding the ecological mechanisms of landscape-wide effects is particularly important for the species groups of edge and farmyard birds, because these birds do not breed in farmland as such but are affected by agricultural land-use as complementary or supplementary habitats (e.g., for foraging or displaying) (Redlich et al. 2018; Smith et al. 2014; Vepsäläinen et al. 2010). In our study, such complementary effects are exemplified for instance by the positive associations between density of edge species and set asides, or density of farmyard bird and areas with leys and pastures in the study landscapes. In addition, we demonstrate that farmyards with animals strongly benefit farmyard bird species, underscoring the importance of mixed production systems at landscape scales. Notably, our results show that grasslands – including leys and pastures as well as subsidized grasslands (which mainly include different kinds of environmental fallows) – benefit open field, FBI, and farmyard species. Thus, interventions targeting nature conservation under the Common Agricultural Policy have the potential to maintain or even increase farmland biodiversity in mosaic landscapes (Hertzog et al. 2023). Yet the observed patterns are likely specific to the landscape context of this study and other Northern European regions, i.e. mosaics of farmland with little winter crop** surrounded by forest.
Land-use and interventions on farmed land
Density of open field, FBI, and farmyard birds increased with larger area of leys and pastures, whereas density of edge species increased with higher patch density of set asides; that is, when the amount and proximity of individual set asides was relatively high within the study landscape, and larger distances between pastures. Density of FBI species, and meadow pipit and tree sparrow abundances increased with larger area of subsidized grasslands, which in our study included environmental fallows intended to promote farmland biodiversity. These results partly corroborate earlier studies showing beneficial effects of extensive grasslands on farmland biodiversity (Herzon et al. 2011; Hertzog et al. 2023; Staggenborg & Anthes 2022; Traba & Morales 2019). Importantly, our results also show that pastures, and to some extent set asides (mainly comprised by stubbles left idle after the previous growing season) provide complementary benefits over and above the provisioning of breeding habitat for species that nest in open farmland. This likely occurs by increasing foraging habitat for species breeding in non-arable habitats (Schmidt et al. 2008). Among individual species, landscape complementation effects can explain why edge and farmyard species (fieldfare, barn swallow, tree sparrow) benefited from leys and pastures.
In a similar way, the positive effect of greater areas of spring-sown crops on total species density could be explained by landscape complementation effects, as some of these bird species forage in arable fields with sparse vegetation during the early breeding season (Devereux et al. 2004; Perkins et al. 2000). However, because of inherent correlations, it is also possible that the effect of greater area of spring-sown crops is driven by the total farmland area within the study landscapes (see SI Fig. S1), suggesting area-abundance mechanisms rather than complementation effects. Furthermore, the effect may also be driven by unaccounted geographical effects. This is likely the case concerning the negative effect of spring-sown cereals on corncrake abundance. The area of spring-sown crops is highest in the southern parts of our study area, where corncrakes are most abundant, while there, the area of grasslands (leys, subsidized grasslands) that the species prefers (Brambilla et al. 2021) is low.
Finally, our results show no effects of larger areas of autumn-sown crops, which is consistent with studies in similar contexts where the proportion of autumn-sown crops is low (Hiron et al. 2013a; Tschumi et al. 2020), but stand in contrast to regions where autumn-sown crops dominate and are mainly associated with declining bird densities (Eggers et al. 2011).
Effects of farmland structures and non-crop habitats
We found that the density of farmyard bird species was up to twice as high in landscapes with larger areas of farmyards with animal husbandry. This effect was stronger than the complementary effects of farmland use described above (see Fig. 2c). The positive effect of farmyards with animals has been previously demonstrated at the local scale, with higher bird species richness and abundance around active (animal) farmyards, in contrast to farmyards with abandoned animal husbandry (Hiron et al. 2013b; Šálek et al. 2018). Considering individual farmyard bird species, we found that tree sparrow and house martin abundances were strongly enhanced by larger areas of farmyards with animal husbandry, which is consistent with the notion that birds with varying insect diets (ground-living and aerial, respectively) benefit from increased food availability on farms with animal husbandry (Santangeli et al. 2019; Tiainen et al. 1989).
Furthermore, a larger area of barns in the study landscapes had a positive association with the abundance of tree sparrows amongst the species-specific responses. This effect of barns and farmyards can be interpreted in two non-exclusive ways. First, barns and farmyards may provide suitable structures and cavities for nesting, display sites, shelter or even foraging on arthropod prey (Rosin et al. 2016). Second, the area of barns and farmyards correlated with the area of farmyards with animals, and might therefore serve as a proxy for study landscapes where mixed farming is still relatively prevalent, capturing this variability in another way compared to the area of farmyards with animals (cf. Benton et al. 2003).
Among non-crop habitats, it is surprising that our landscape-level analysis did not reveal stronger effects of ditches on bird assemblages, given that high density of non-crop field borders has been shown to be more important for farmland birds than in-field interventions in local-scale studies conducted partly in the same study region (Ekroos et al. 2019). Ditches are beneficial landscape features in farmlands, as they support various aquatic and terrestrial taxa, provide food resources and serve as connecting corridors in the landscape (Herzon & Helenius 2008); while ditch verges with a more complex vegetation structure are particularly important (Marja & Herzon 2012). In this study, we show that whinchat abundance almost doubled with greater relative ditch areas, most likely because they use the ditch verges as nesting sites. However, the density of FBI species was lower in landscapes with more ditches. Low density of FBI species in landscapes with extremely high ditch verge cover (> 50%) were most likely driven by two of the northern-most study landscapes, which had very large extents of ditch verges and, because of their higher latitude, overall bird densities were low compared to southern study landscapes (SI Fig. S22).
Effects of landscape mosaic variables
One main finding of this study is that the total density of all species was strongly correlated with greater habitat Shannon diversity, as well as the strong positive link between habitat richness and density of FBI species. At the species level, we show that jackdaws and barn swallows – both species of the Finnish FBI – benefitted from a greater habitat Shannon diversity and greater richness of farmland types, respectively. The positive effect of the latter, which predominantly reflects the richness of crop types and farmland structures in the landscape, contrasts with earlier studies that focused on crop diversity at smaller spatial scales (Ekroos et al. 2019; Josefsson et al. 2017; McDaniel et al. 2014; Palmu et al. 2014). Our results suggest that a higher diversity of crops and mixed farming benefit some of the species selected to monitor the state of farmland birds. On the other hand, lapwings were negatively associated with habitat richness, likely reflecting the species’ specialization on open fields as their primary habitat (i.e., positive effect of leys and pastures), which reflects the general idea that ecological specialists are expected to benefit from more homogeneous environments as compared to generalists relying on more heterogeneous landscapes (Östergård & Ehrlén 2005).
Density of open field birds consistently increased with greater area of farmland in the study landscapes, whereas total bird density and density of edge species decreased with greater areas of farmed land. In boreal regions, farmland is typically organized as patches mainly surrounded by forests, leading to an area-abundance relationship for birds mainly dependent on farmland habitats such as open field species (cf. Vepsäläinen et al. 2010). This is reflected also in the individual species responses where ortolan buntings were strongly positively associated with greater farmland area. However, the density of edge species and total species did not show the same relationship, as their densities are likely influenced by the proximity to farmland structures such as farmyards and an overall higher habitat heterogeneity at the landscape scale.
Additional species-specific responses
In addition to the effects on individual species discussed above, we identified several important additional habitat associations at the species level. First, we found weak effects of clear-cuts on farmland bird abundances (i.e., skylark and whitethroat), and all significant effects were negative (Table 4). This finding was somewhat surprising, as some open farmland bird species benefit from clearcuts (Ram et al. 2020). Skylarks seem to avoid clear-cuts, whereas whinchats prefer them in a regional comparison of habitat types (Ram et al. 2020). Because our bird surveys did not comprehensively include clearcuts (except for their edges which were monitored if they were situated next to farmland), the negative association between clearcuts and whitethroats could potentially be driven by a dilution effect, as whitethroats may use clearcuts as well (Ram et al. 2020). In the case of skylarks, they may avoid clearcuts because of a lack of nesting sites (Ram et al. 2020), or a proximity to forest edges which skylarks tend to avoid (Ekroos et al. 2019; Piha et al. 2003). As our bird censuses did not explicitly cover clearcuts, their relative importance and ecological mechanisms affecting farmland birds in mosaic landscapes need to be confirmed by additional research.
Second, we found effects of a larger-scale landscape context, with whinchat and whitethroat abundances decreasing with larger distances between forest patches within a radius of 5 km, and lapwing, curlew and jackdaw abundances decreasing with larger distances between pastures (lapwings) or arable land (curlews and jackdaws). Lapwings, curlews and jackdaws form loose colonies or territory groups where they are abundant. Because none of the three species occupy home-ranges across such large extents, the result is most likely driven by source-sink dynamics (Pulliam 1988; Smith et al. 2014), added with social attraction dynamics (e.g., Oh & Badyaev 2010), such that areas situated in regions with a high proportion of arable land and pastures maintain viable source populations. The preference for shorter distances between forest patches exhibited by whinchats and whitethroats likely demonstrates that they benefit from a configurationally heterogeneous environment in which farmland is interspersed with forest patches, which in turn translates into a higher share of forest edge habitats (Berg & Pärt 1994).
Finally, skylark abundance increased with area of direct-sown spring crops. Fields with direct-sown spring crops in our study are comparable with reduced tilling, or no-till farming systems, the latter of which is positively associated with nest densities of farmland birds (VanBeek et al. 2014). As direct-sown spring crop fields are not ploughed, they likely provide seed and insect food early in the season (c.f. Cunningham et al. 2004), and because tillage and sowing are done early, skylark nests are likely to be relatively safe from damage caused by farming operations (c.f. VanBeek et al. 2014).
Conclusions
With our landscape-scale approach, we show effects of various land-use interventions that are likely either due to direct habitat effects (e.g., positive effect of farmyards with animals on farmyard bird species), or because of higher availability of multiple habitats providing complementary resources. The latter explanation is based on two observations. First, we found that agricultural land-use matters for species groups that forage in farmland but do not breed in farmland habitats, and thus rely on multiple habitat types (e.g., farmyard species benefit from leys and pastures). Second, we show positive effects of habitat Shannon diversity – a common measure for landscape heterogeneity– as well as habitat richness on different bird species and species guilds, which is consistent with effects caused by landscape and resource complementation. Our findings demonstrate that farmland interventions such as leys and pastures, set asides, or subsidized grasslands support higher densities of farmland birds, confirming that extensive grasslands and crop rotation systems support higher biodiversity and thus should be implemented by practitioners and supported through farmland policies such as the Common Agricultural Policy or EU’s Nature Restoration Law (https://environment.ec.europa.eu/topics/nature-and-biodiversity/nature-restoration-law_en) to ensure their implementation also in the future. Notably, we demonstrate that in addition to farmland interventions as such, mixed farming (i.e., crops and animal farms), as well as farmland structures such as farmyards or barns strongly contribute to higher bird densities in the landscape. Although the number of farms with animal husbandry is projected to decrease in Finland (Lehtonen et al. 2020), our results suggest that it will be important to find incentives to maintain animal husbandry, particularly in regions dominated by cereal farming. From a compositional viewpoint, overall landscape heterogeneity supports higher bird numbers. This is especially important for FBI species, which are currently used as an indicator of farmland biodiversity more broadly. Hence, this should be promoted also in the future through, for example, financial support to small-scale farms that employ extensive land-use management practices and are not specialized in only one or a few crop types.
Data availability
Data and R codes are available upon request from the authors.
References
Batáry P, Matthiesen T, Tscharntke T (2010) Landscape-moderated importance of hedges in conserving farmland bird diversity of organic vs. conventional croplands and grasslands. Biol Conserv 143:2020–2027
Batáry P, Dicks LV, Kleijn D, Sutherland WJ (2015) The role of agri-environment schemes in conservation and environmental management. Conserv Biol 29:1006–1016
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Berg Å, Pärt T (1994) Abundance of breeding farmland birds on arable and set-aside fields at forest edges. Ecography 17(2):147–152
Bianchi FJJA, Booij CJH, Tscharntke T (2006) Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proc R Soc B 273:1715–1727
Brambilla M, Gubert F, Pedrini P (2021) The effects of farming intensification on an iconic grassland bird species, or why mountain refuges no longer work for farmland biodiversity. Agric Ecosyst Environ 319:107518. https://doi.org/10.1016/j.agee.2021.107518
Brlík V, Šilarová E, Škorpilová J, Alonso H, Anton M, Aunins A et al (2021) Long-term and large-scale multispecies dataset tracking population changes of common European breeding birds. Sci Data 8:21
Bruun M, Smith HG (2003) Landscape composition affects habitat use and foraging flight distances in breeding European starlings. Biol Conserv 114:179–187
Cook RD (1977) Detection of Influential Observation in Linear Regression. Technometrics 19(1):15–18. https://doi.org/10.1080/00401706.1977.10489493
Cunningham HM, Chaney K, Bradbury RB, Wilcox A (2004) Non-inversion tillage and farmland birds: a review with special reference to the UK and Europe. Ibis 146:192–202. https://doi.org/10.1111/j.1474-919X.2004.00354.x
Devereux CL, McKeever CU, Benton TG, Whittingham MJ (2004) The effect of sward height and drainage on Common Starlings Sturnus vulgaris and Northern Lapwings Vanellus vanellus foraging in grassland habitats. Ibis 146:115–122
Donald PF, Green RE, Heath MF, Donal PF, Gree RE, Heath MF (2001) Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc R Soc B 268:25–29
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G et al (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46
EEA (2013). Annual report 2012 and Environmental statement 2013. Copenhagen: European Environment Agency. 95 pp. https://doi.org/10.2800/91164
Eggers S, Unell M, Pärt T (2011) Autumn-sowing of cereals reduces breeding bird numbers in a heterogeneous agricultural landscape. Biol Conserv 144:1137–1144
Eglington SM, Pearce-Higgins JW (2012) Disentangling the Relative Importance of Changes in Climate and Land-Use Intensity in Driving Recent Bird Population Trends. PLoS ONE 7:e30407
Ekroos J, Tiainen J, Seimola T, Herzon I (2019) Weak effects of farming practices corresponding to agricultural greening measures on farmland bird diversity in boreal landscapes. Landsc Ecol 34:389–402
Eurostat. (2023). Animal production statistics. https://ec.europa.eu/eurostat/databrowser/view/AGR_R_ANIMAL/default/line?lang=en
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ et al (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14(2):101–112. https://doi.org/10.1111/j.1461-0248.2010.01559x
Gregory RD, Van Strien AJ, Vorisek P, Meyling AWGG, Noble DG, Foppen RPBB et al (2005) Develo** indicators for European birds. Phil Trans R Soc B 360:269–288
Grüebler MU, Korner-Nievergelt F, von Hirschheydt J (2010) The reproductive benefits of livestock farming in barn swallows Hirundo rustica: quality of nest site or foraging habitat? J Appl Ecol 47:1340–1347
GTK. (2018). Superficial deposits of Finland 1:200 000 (sediment polygons). Geologian tutkimuskeskus GTK. http://tupa.gtk.fi/raportti/arkisto/m10_4_2010_3.pdf & https://hakku.gtk.fi/
Hertzog LR, Klimek S, Röder N, Frank C, Böhner HG, Kamp J (2023) Associations between farmland birds and fallow area at large scales: Consistently positive over three periods of the EU Common Agricultural Policy but moderated by landscape complexity. J Appl Ecol 60(6):1077–1088. https://doi.org/10.1111/1365-2664.14400
Herzon I, Helenius J (2008) Agricultural drainage ditches, their biological importance and functioning. Biol Conserv 141:1171–1183
Herzon I, Ekroos J, Rintala J, Tiainen J, Seimola T, Vepsäläinen V (2011) Importance of set-aside for breeding birds of open farmland in Finland. Agric Ecosyst Environ 143:3–7
Hesselbarth MH, Sciaini M, With KA, Wiegand K, Nowosad J (2019) Landscapemetrics: an open‐source R tool to calculate landscape metrics. Ecography 42(10):1648–1657
Hiron M, Berg Å, Eggers S, Josefsson J, Pärt T (2013a) Bird diversity relates to agri-environment schemes at local and landscape level in intensive farmland. Agric Ecosys Environ 176:9–16
Hiron M, Berg Å, Eggers S, Pärt T (2013b) Are farmsteads over-looked biodiversity hotspots in intensive agricultural ecosystems? Biol Conserv 159:332–342
Hiron M, Berg Å, Eggers S, Berggren Å, Josefsson J, Pärt T (2015) The relationship of bird diversity to crop and non-crop heterogeneity in agricultural landscapes. Landsc Ecol 30:2001–2013
Josefsson J, Berg Å, Hiron M, Pärt T, Eggers S (2017) Sensitivity of the farmland bird community to crop diversification in Sweden: does the CAP fit? J Appl Ecol 54:518–526. https://doi.org/10.1111/1365-2664.12779
Kleijn D, Rundlöf M, Scheper J, Smith HG, Tscharntke T (2011) Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol Evol 26:474–481
Lehtonen H, Saarnio S, Rantala J, Luostarinen S, Maanavilja L, Heikkinen J, Soini K, Aakkula J, Jallinoja M, Rasi S, Niemi J (2020) Maatalouden ilmastotiekartta – Tiekartta kasvihuonekaasupäästöjen vähentämiseen Suomen maataloudessa. Maa- ja metsätaloustuottajain Keskusliitto MTK ry. Helsinki
Leys C, Ley C, Klein O, Bernard P, Licata L (2013) Detecting outliers: Do not use standard deviation around the mean, use absolute deviation around the median. J Experim Soc Psychol 49(4):764–766
Lüdecke D, Ben-Shachar M, Patil I, Waggoner P, Makowski D (2021) performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J Open Source Softw 6:3139
Marja R, Herzon I (2012) The importance of drainage ditches for farmland birds in agricultural landscapes in the Baltic countries: does field type matter? Ornis Fenn 89:170–181
Maxwell D, Robinson DA, Thomas A, Jackson B, Maskell L, Jones DL et al (2017) Potential contribution of soil diversity and abundance metrics to identifying high nature value farmland (HNV). Geoderma 305:417–432
McDaniel MD, Tiemann LK, Grandy AS (2014) Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol Appl 24:560–570
Meier ES, Lüscher G, Knop E (2022) Disentangling direct and indirect drivers of farmland biodiversity at landscape scale. Ecol Lett 25:2422–2434
Newton I (2017) Farming and birds (Collins New Naturalist Library, Book 135) (Vol 135). HarperCollins UK
Official Statistics of Finland (2022) Structure of agricultural and horticultural enterprises 2022 [web publication]. Helsinki: Natural Resources Institute Finland [referred: 13.3.2024]. Access method: https://www.luke.fi/en/statistics/structure-of-agricultural-and-horticultural-enterprises/structure-of-agricultural-and-horticultural-enterprises-2022
Oh KP, Badyaev AP (2010) Structure of social networks in passerine birds: consequences for sexual selection and the evolution of mating strategies. Am Nat 176:E80–E89
Östergård H, Ehrlén J (2005) Among population variation in specialist and generalist seed predation–the importance of host plant distribution, alternative hosts and environmental variation. Oikos 111(1):39–46
Palmu E, Ekroos J, Hanson HI, Smith HG, Hedlund K (2014) Landscape-scale crop diversity interacts with local management to determine ground beetle diversity. Basic Appl Ecol 15:241–249
Pe’er G, Zinngrebe Y, Hauck J, Schindler S, Dittrich A, Zingg S (2017) Adding Some Green to the Greening: Improving the EU’s Ecological Focus Areas for Biodiversity and Farmers. Conserv Lett 10:517–530
PECBMS. (2021a). Common farmland birds indicator Europe 1980–2021. EBCC/BirdLife/RSPB/CSO: https://pecbms.info/trends-and-indicators/indicators/all/yes/indicators/E_C_Fa/
PECBMS. (2021b). Common forest birds indicator Europe 1980–2021. EBCC/BirdLife/RSPB/CSO: https://pecbms.info/trends-and-indicators/indicators/all/yes/indicators/E_C_Fo/
Perkins AJ, Whittingham MJ, Bradbury RB, Wilson JD, Morris AJ, Barnett PR (2000) Habitat characteristics affecting use of lowland agricultural grassland by birds in winter. Biol Conserv 95:279–294
Piha M, Pakkala T, Tiainen J (2003) Habitat preferences of the Skylark Alauda arvensis in southern Finland. Ornis Fenn 80:97–110
Pulliam HR (1988) Sources, sinks, and population regulation. Am Nat 132:652–661
R Core Team. (2022). A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/. R version 4.2.0 (2022–04–22 ucrt) - "Vigorous Calisthenics"
Ram D, Lindström Å, Pettersson LB, Caplat P (2020). Forest clear-cuts as habitat for farmland birds and butterflies. For Ecol Manage, 473. https://doi.org/10.1016/j.foreco.2020.118239
Redlich S, Martin EA, Wende B, Steffan-Dewenter I (2018) Landscape heterogeneity rather than crop diversity mediates bird diversity in agricultural landscapes. PLoS ONE 13:e0200438
Rigal S, Dakos V, Alonso H, Auniņš A, Benkő Z, Brotons L, Devictor V (2023) Farmland practices are driving bird population decline across Europe. Proc Natl Acad Sci 120(21):e2216573120
Robinson RA, Sutherland WJ (2002) Post-war changes in arable farming and biodiversity in Great Britain. J Appl Ecol 39:157–176
Rosin ZM, Skorka P, Part T, Zmihorski M, Ekner-Grzyb A, Kwiecinski Z et al (2016) Villages and their old farmsteads are hot spots of bird diversity in agricultural landscapes. J Appl Ecol 53:1363–1372
Šálek M, Bazant M, Zmihorski M (2018) Active farmsteads are year-round strongholds for farmland birds. J Appl Ecol 55:1908–1918
Šálek M, Brlík V, Kadava L, Praus L, Studecký J, Vrána J et al (2020) Year-round relevance of manure heaps and its conservation potential for declining farmland birds in agricultural landscape. Agric Ecosyst Environ 301:107032
Santangeli A, Lehikoinen A, Lindholm T, Lindholm T, Herzon I (2019) Organic animal farms increase farmland bird abundance in the Boreal region. PLoS ONE 14:1–16
Schmidt MH, Rocker S, Hanafi J, Gigon A (2008) Rotational fallows as overwintering habitat for grassland arthropods: the case of spiders in fen meadows. Biodivers Conserv 17:3003–3012
Siriwardena GM (2010) The importance of spatial and temporal scale for agri-environment scheme delivery. Ibis 152:515–529
Smith HG, Birkhofer K, Clough Y, Ekroos J, Olsson O, Rundlöf M (2014) Beyond dispersal: the role of animal movement in modern agricultural landscapes. In: Animal Movement Across Scales. Oxford University Press, pp 51–70
Staggenborg J, Anthes N (2022) Long‐term fallows rate best among agri‐environment scheme effects on farmland birds – A meta‐analysis. Conserv Lett 15(4):e12904
Stjernman M, Sahlin U, Olsson O, Smith HG (2019) Estimating effects of arable land use intensity on farmland birds using joint species modeling. Ecol Appl 29:e01875
SYKE (2012) CORINE Land Cover 2012, 20x20m. Suomen ympäristökeskus SYKE. https://ckan.ymparisto.fi/dataset/corine-maanpeite-2012
Tiainen J, Hanski IK, Pakkala T, Piiroinen J, Yrjölä R (1989) Clutch size, nestling growth and nestling mortality of the Starling Sturnus vulgaris in south Finnish agroenvironments. Ornis Fenn 66:41–48
Tiainen J, Pakkala T (2001) Birds. In: Pitkänen M, Tiainen J (eds) Biodiversity in agricultural landscapes in Finland. BirdLife Finland Conservation Series 3:33–50
Traba J, Morales MB (2019) The decline of farmland birds in Spain is strongly associated to the loss of fallowland. Sci Rep 9:9473
Tscharntke T, Tylianakis JM, Rand TA, Didham RK, Fahrig L, Batáry P et al (2012) Landscape moderation of biodiversity patterns and processes - eight hypotheses. Biol Rev 87:661–685
Tschumi M, Birkhofer K, Blasiusson S, Jorgensen M, Smith HG, Ekroos J (2020) Woody elements benefit bird diversity to a larger extent than semi-natural grasslands in cereal-dominated landscapes. Basic Appl Ecol 46:15–23
VanBeek KR, Brawn JD, Ward MP (2014) Does no-till soybean farming provide any benefits for birds? Agric Ecosyst Environ 185:59–64. https://doi.org/10.1016/j.agee.2013.12.007
Venables WN, Ripley BD (2002) Modern applied statistics with S, 4th edn. Springer, New York
Vepsäläinen V, Tiainen J, Holopainen J, Piha M, Seimola T (2010) Improvements in the Finnish agri-environment scheme are needed in order to support rich farmland avifauna. Ann Zool Fennici 47:287–305
Vickery J, Arlettaz R (2012) The importance of habitat heterogeneity at multiple scales for birds in European agricultural landscapes. In: Fuller RJ (ed) Birds and Habitat: Relationships in Changing Landscapes. Cambridge University Press, Cambridge, pp 177–204
Vickery JA, Feber RE, Fuller RJ (2009) Arable field margins managed for biodiversity conservation: A review of food resource provision for farmland birds. Agric Ecosyst Environ 133:1–13
Zingg S, Ritschard E, Arlettaz R, Humbert J-Y (2019) Increasing the proportion and quality of land under agri-environment schemes promotes birds and butterflies at the landscape scale. Biol Conserv 231:39–48
Żmihorski M, Berg Å, Pärt T (2016) Forest clear-cuts as additional habitat for breeding farmland birds in crisis. Agric Ecosyst Environ 233:291–297
Acknowledgements
We thank all farmers that became engaged with this project, Hannu Hiisivuori, Sampo Laukkanen, Kalle Meller, and Jarmo Piiroinen for assistance in bird monitoring. We also thank Gavin Siriwardena and an anonymous reviewer for constructive comments on an earlier draft of this study.
Funding
Open Access funding provided by University of Helsinki (including Helsinki University Central Hospital). The OPAL Life-project, and the Ministry of Agriculture and Forestry provided funding to JT, TS and LB. In addition, AL received funding from Research Council of Finland (grant 323527), and LB from Kone Foundation (grant Nr. 202104881).
Author information
Authors and Affiliations
Contributions
Juha Tiainen, Tuomas Seimola, Markus Piha, and Johan Ekroos contributed to the study conception and design. Juha Tiainen and Tuomas Seimola conducted field work while Tuomas Seimola digitized the farmland habitat map**. Laura Bosco and Johan Ekroos analysed the data. The first draft of the manuscript was written by Laura Bosco and Johan Ekroos and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bosco, L., Lehikoinen, A., Piha, M. et al. Relative effects of arable land-use, farming system and agri-environment schemes on landscape-scale farmland bird assemblages. Landsc Ecol 39, 113 (2024). https://doi.org/10.1007/s10980-024-01906-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01906-z