
Abstract
Background
There is a great demand for antibiotic alternatives to maintain animal health and productivity. The objective of this experiment was to determine the efficacy of dietary supplementation of a blood group A6 type 1 antigen oligosaccharides-based polymer (Coligo) on growth performance, diarrhea severity, intestinal health, and systemic immunity of weaned pigs experimentally infected with an enterotoxigenic Escherichia coli (ETEC), when compared with antibiotics.
Results
Pigs in antibiotic carbadox or Coligo treatment groups had greater (P < 0.05) body weight on d 5 or d 11 post-inoculation (PI) than pigs in the control group, respectively. Supplementation of antibiotics or Coligo enhanced (P < 0.05) feed efficiency from d 0 to 5 PI and reduced (P < 0.05) frequency of diarrhea throughout the experiment, compared with pigs in the control group. Supplementation of antibiotics reduced (P < 0.05) fecal β-hemolytic coliforms on d 2, 5, and 8 PI. Pigs in antibiotics or Coligo groups had reduced (P < 0.05) neutrophil counts and serum haptoglobin concentration compared to pigs in the control group on d 2 and 5 PI. Pigs in Coligo had reduced (P < 0.05) total coliforms in mesenteric lymph nodes on d 5 and 11 PI, whereas pigs in antibiotics or Coligo groups had reduced (P < 0.05) total coliforms in spleen on d 11 PI compared with pigs in the control group. On d 5 PI, pigs in the Coligo group had greater (P < 0.05) gene expression of ZO1 in jejunal mucosa, but less (P < 0.05) mRNA expression of IL1B, IL6, and TNF in ileal mucosa, in comparison with pigs in the control group. Supplementation of antibiotics enhanced (P < 0.05) the gene expression of OCLN in jejunal mucosa but decreased (P < 0.05) IL1B and IL6 gene expression in ileal mucosa, compared with the control. On d 11 PI, supplementation of antibiotics or Coligo up-regulated (P < 0.05) gene expression of CLDN1 in jejunal mucosa, but Coligo reduced (P < 0.05) IL6 gene expression in ileal mucosa compared to pigs in the control group.
Conclusions
Supplementation of Coligo improved growth performance, alleviated diarrhea severity, and enhanced gut health in weaned pigs infected with ETEC F18 in a manner similar to in-feed antibiotics.
Similar content being viewed by others

Background
Enterotoxigenic E. coli (ETEC) strains expressing F4 or F18 fimbriae are major causes of post-weaning diarrhea in nursery pigs [1]. Attachment of ETEC to the specific receptors on intestinal epithelium leads to colonization and secretion of enterotoxins, resulting in secretory diarrhea in weanling pigs [2]. To prevent post-weaning diarrhea and improve production of pigs, antibiotics were commonly added to the diet over the past decades. However, frequent use of in-feed antibiotics in livestock production has been shown to contribute to the increased prevalence of antibiotic-resistant bacteria and raised public health concerns [3]. With these issues, the Food and Drug Administration (FDA) banned the use of in-feed antibiotics for growth promoting purposes in livestock production in the U.S. [4], thus alternative nutritional strategies are highly demanded to enhance disease resistance and production of weanling pigs.
Many nutritional approaches have been applied to prevent post-weaning diarrhea associated with ETEC and enhance the production of pigs. Among these, the prevention of bacterial attachment to the small intestine is one of the most effective defense strategies against ETEC infection [5]. Oligosaccharides have been reported to possess ETEC receptor activity for bacterial adhesions [6, 7]. Especially, N-acetylgalactosamine (GalNAc) containing glycans that could enhance the binding affinity of various ETEC strains, including K99 [8], F4 [9], and F18 [10,11,12]. Coddens et al. [11] identified the blood group H type 1 determinant (Fuca2Galß3GlcNAc) as the minimal binding epitope of F18 fimbriae. Based on that, an optimal binding epitope was created by adding the terminal 3-linked galactose or N-acetylgalactosamine of the blood group B type 1 determinant (Galα3(Fucα2)Galß3GlcNAc) and the blood group A type 1 determinant (GalNAcα3(Fucα2)- Galß3GlcNAc). The purified soluble blood group oligosaccharides were able to greatly reduce binding of F18 positive E. coli to intestinal villi of F18 receptor-positive pigs in concentrations of 1 to 10 mg/mL [11]. Moreover, multimerizing blood group A on human serum albumin reduced the amount of blood group oligosaccharides needed 1000 times [13]. Therefore, multimerizing the blood group A oligosaccharides may efficiently prevent enterotoxin-induced secretory diarrhea. Recently, grafted polymers that combine multiple substances have been proposed for their potential synergistic effects on preventing human and animal diseases [14, 15]. Epsilon-poly-lysin (ε-PL) has been reported to use as a carrier in the membrane transport of proteins and drugs [16]. Due to its excellent heat stability, biodegradability, and lack of toxicity, ε-PL has generally been regarded as safe status (GRAS) and been interested in the food and medicine industries as a delivery vehicle targeting the desired location [17]. Thus, the combination of blood group oligosaccharides and ε-PL may enhance the resistance of pigs against F18 ETEC infection by inhibiting bacterial attachment and/or directly killing bacteria. The overall objective of this study was to investigate the efficacy of blood group A6 type 1-based polymer on gut integrity and disease resistance of weanling pigs challenged with F18 ETEC.
Materials and methods
Animals, housing, experimental design, and diet
A total of 48 weanling pigs (crossbred; initial body weight (BW): 7.23 ± 1.14 kg; 21 days old) with an equal number of gilts and barrows were used in this study. They were selected from the Swine Teaching and Research Center at the University of California, Davis. The sows and piglets used in this experiment did not receive E. coli vaccines, antibiotic injections, or antibiotics in creep feed. Before weaning, feces were collected from sows and all their piglets destined for this study to verify the absence of β-hemolytic E. coli. The F18 ETEC receptor status was also tested based on the methods of Kreuzer et al. [18], and all piglets used in this study were susceptible to F18 ETEC.
After weaning, all pigs were randomly assigned to one of the four dietary treatments in a randomized complete block design with body weight within sex and litter as the blocks and pig as the experimental unit. There were 12 replicate pigs per treatment. Pigs were individually housed (pen size: 0.61m × 1.22 m) in environmental control rooms at the Cole Facility at the University of California, Davis for 18 d, including 7 d before and 11 d after the first F18 ETEC challenge (d 0). The piglets had ad libitum access to feed and water. Environmental enrichment was provided for each pig. The light was on at 07:00 h and off at 19:00 h daily in the environmental control rooms.
The 4 dietary treatments included: 1) Positive control: control diet; 2) Low dose oligosaccharide-based polymer (LOW): control diet supplemented with 10 mg/kg oligosaccharide-based polymer active substance (Coligo); 3) High dose oligosaccharide-based polymer (HIGH): control diet supplemented with 20 mg/kg oligosaccharide-based polymer active substance (Coligo); and 4) CAR: control diet supplemented with 50 mg/kg carbadox. Spray-dried plasma and high levels of zinc oxide exceeding recommendation and normal practice were not included in the diets. The experimental diets were fed to pigs throughout the experiment. Oligosaccharide-based polymer active substance was a glycoconjugate composed of blood group A6 type 1 antigen oligosaccharides grafted on a single peptide of epsilon-poly-lysine. Coligo was designed and synthesized by Elicityl (France) in cooperation with Ghent University (Dr. Eric Cox’s laboratory) and was provided by Pancosma (Geneva, Switzerland). The mean rate of conjugation is 15 mol of oligosaccharide for 1 mol of ε-PL. The oligosaccharide part represents 25% of the molecular weight of the oligosaccharide-based polymer. All diets were formulated to meet pig nutritional requirements (Table 1) [19] and provided as mash form throughout the experiment.
After 7 days of adaptation, all pigs were orally inoculated with 3 mL of F18 ETEC for 3 consecutive days from d 0 post-inoculation (PI). The F18 ETEC was originally isolated from a field disease outbreak by the University of Illinois Veterinary Diagnostic Lab (isolate number: U.IL-VDL # 05–27,242). The F18 ETEC expresses heat-labile toxin (LT), heat-stable toxin b (STb), and Shiga-like toxin (Stx2e). The inoculums were prepared by the laboratory of the Western Institute for Food Safety and Security at the University of California, Davis, and were provided at 1010 colony-forming unit (CFU) per 3 mL dose in phosphate-buffered saline (PBS). This dose caused mild diarrhea in the current study, consistent with our previous published researches [20,21,1). The qRT-PCR reaction conditions followed the published research [27]. The 2-ΔΔCT method was used to analyze the relative expression of genes compared with control [28].
Statistical analysis
The normality of data was verified with the Shapiro-Wilk test, and outliers were identified using the UNIVARIATE procedure (SAS Inst. Inc., Cary, NC, USA). All data were analyzed by ANOVA using the PROC MIXED of SAS (SAS Institute Inc., Cary, NC, USA) in a randomized complete block design with the pig as the experimental unit. The statistical model included independent variables treatment group, sampling day, and interactions as the fixed effect and blocks as random effects. Treatment means were separated by using the LSMEANS statement and the PDIFF option of PROC MIXED. Contrast statements were used to analyze the dose effects of Coligo. The Chi-square test was used for analyzing the frequency of diarrhea. Statistical significance and tendency were considered at P < 0.05 and 0.05 ≤ P < 0.10, respectively.
Results
Growth performance, diarrhea score, β-hemolytic coliforms
No difference was observed in the initial BW and d 0 BW of pigs among dietary treatments (Table 2). Pigs supplemented with CAR had greater (P < 0.05) BW on d 5 PI than pigs in the control and HIGH groups. Pigs supplemented with LOW had the greatest (P < 0.05) BW, but pigs supplemented with HIGH had the lowest (P < 0.05) BW on d 11 PI among all dietary treatments. Supplementation of LOW had greater (P < 0.05) ADFI of pigs from d 5 to 11 PI, compared with control and HIGH groups. Supplementation of Coligo had greater (P < 0.05) feed efficiency from d 0 to 5 PI compared with pigs in the control group regardless of dose. Supplementation of HIGH also had greater (P < 0.05) feed efficiency of weaned pigs from d 5 to 11 PI, compared with pigs in the control. Pigs fed with CAR had better (P < 0.05) feed efficiency than pigs fed with the control diet from d 0 to 5 PI, but this was not the case from d 5 to 11 PI.
Pigs supplemented with CAR had the lowest (P < 0.05) average diarrhea score from d 0 to 5 PI and d 5 to 11 PI among all dietary treatments (Table 3; Fig. 1). Compared with pigs in control group, pigs supplemented with LOW had lower (P < 0.05) average diarrhea score of weaned pigs from d 0 to 5 PI, but this was not the case from d 5 to 11 PI. Supplementation of CAR or any dose of Coligo had lower (P < 0.05) frequency of diarrhea of weaned pigs from d 0 to 11 PI.

Daily diarrhea score of ETEC-infected weaned pigs fed diets supplemented with oligosaccharide-based polymer (Coligo) or antibiotics. Diarrhea score = 1, normal feces, 2, moist feces, 3, mild diarrhea, 4, severe diarrhea, 5, watery diarrhea. Each least squares mean from d 0 to d 5 post-inoculation (PI) represents 12 observations. Each least squares mean from d 6 to d 11 PI represents 6 observations. *Significant differences were observed among dietary treatment: P < 0.05. LOW = Low dose blood group A6 type 1-based polymer (Coligo); HIGH = High dose blood group A6 type 1-based polymer (Coligo); CAR = Carbadox
No β-hemolytic coliform was observed in the feces of all pigs before ETEC inoculation. Pigs supplemented with CAR had the lowest (P < 0.05) β-hemolytic coliform percentage in feces on d 2 and 5 PI among all dietary treatments (Fig. 2). The percentage of β-hemolytic coliform in feces was not different between Coligo groups and CAR on d 8 PI. There were no differences observed in fecal culture on d 11 PI among the treatments.

The percentage (%) of β-hemolytic coliform in fecal samples of ETEC-infected pigs fed diets supplemented with oligosaccharide-based polymer (Coligo) or antibiotics. Each least squares mean from d 0 to d 5 post-inoculation (PI) represents 12 observations. Each least squares mean from d 6 to d 11 PI represents 6 observations. a,bMeans without a common superscript differ (P < 0.05). LOW = Low dose blood group A6 type 1-based polymer (Coligo); HIGH = High dose blood group A6 type 1-based polymer (Coligo); CAR = Carbadox
Systemic immunity and red blood cell profile
Lymphocyte counts were greater (P < 0.05) in pigs fed CAR on d 0 before ETEC inoculation (Table 4). Pigs in the LOW group had lower (P < 0.05) neutrophils, lymphocytes, and basophils on d 2 PI and lower (P < 0.05) neutrophil counts on d 5 PI, compared with pigs in the control group. Supplementation of HIGH also had lower (P < 0.05) white blood cell counts, neutrophils, lymphocytes, and basophils on d 2 PI. Pigs in the CAR group had lower (P < 0.05) neutrophils and basophils on d 2 PI and lower (P < 0.05) neutrophils on d 5 PI, but higher (P < 0.05) eosinophils on d 5 PI, compared with pigs in control group. No difference was observed in white blood cell profiles among treatments on d 11 PI.
No difference was observed in serum TNF-α concentration among dietary treatments throughout the experiment. Compared with the pigs fed control diet, pigs supplemented with LOW had lower (P < 0.05) haptoglobin on d 5 PI, while pigs fed CAR had lower (P < 0.05) C-reactive protein on d 2, 5, and 11 PI and had lower (P < 0.05) haptoglobin on d 5 PI. No differences in serum C-reactive protein and haptoglobin were observed between the control and HIGH groups.
Before ETEC inoculation, pigs in the CAR group had the lowest (P < 0.05) mean corpuscular volume and total platelets among all dietary treatments on d 0 (Table S2). Supplementation of LOW had lower (P < 0.05) red blood cells and packed cell volume on d 2 PI, while supplementation of HIGH had lower (P < 0.05) packed cell volume on d 5 PI, compared with pigs in the control. Pigs supplemented with CAR had lower (P < 0.05) red blood cells and packed cell volume, but higher (P < 0.05) mean corpuscular hemoglobin and mean corpuscular hemoglobin concentration on d 2 and 5 PI, compared with pigs in the control. Supplementation of CAR also had greater (P < 0.05) total protein concentration on d 11 PI in comparison to pigs in the other treatments.
Bacterial translocation
Supplementation of HIGH had lower (P < 0.05) bacterial translocation in lymph nodes on d 5 and 11 PI compared with control group (Fig. 3). Pigs supplemented with Coligo or CAR had lower (P < 0.05) bacterial translocation in the spleen than pigs in the control on d 11 PI.

Bacterial counts (CFU/g) in lymph node and spleen of ETEC-infected weaned pigs fed diets supplemented with oligosaccharide-based polymer (Coligo) or antibiotics. Each least squares mean from d 0 to d 5 post-inoculation (PI) represents 12 observations. Each least squares mean from d 6 to d 11 PI represents 6 observations. a,bMeans without a common superscript differ (P < 0.05). LOW = Low dose blood group A6 type 1-based polymer (Coligo); HIGH = High dose blood group A6 type 1-based polymer (Coligo); CAR = Carbadox
Intestinal morphology
On d 5 PI, supplementation of Coligo dose-dependently had greater (linear, P < 0.05) villi height, the ratio of villi height to crypt depth, villi width, and villi area in duodenum, had greater (linear, P < 0.05) the ratio of villi height to crypt depth in jejunum, and had greater (linear, P < 0.05) villi height, the ratio of villi height to crypt depth, and villi area in ileum, compared with the control group (Table S3). Supplementation of Coligo also had greater (linear, P < 0.05) duodenal and jejunal villi height and jejunal and ileal villi area, and tended to have greater (linear, P < 0.10) the ratio of villi height to crypt depth in jejunum and ileal villi height on d 11 PI. Pigs fed with CAR had greater (P < 0.05) villi height in duodenum and ileum, the ratio of villi height to crypt depth in all three intestinal segments, and villi area in duodenum than pigs in the control group on d 5 PI. On d 11 PI, pigs supplemented with CAR had higher (P < 0.05) villi height in all three intestinal segments, greater (P < 0.05) villi height to crypt depth ratio in jejunum, and bigger (P < 0.05) sialomucin area in duodenum than pigs in the control group. In addition, pigs in the CAR group also had greater (P < 0.05) villi height:crypt depth in all intestinal segments on d 5 PI, and greater (P < 0.05) villi height in ileum, in comparison to pigs in the Coligo treatment group.
Intestinal barrier and innate immunity
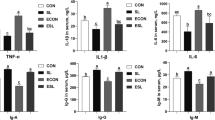
No difference was observed in the mRNA expression of MUC2 in jejunal mucosa among pigs in all dietary treatment groups (Fig.4). On d 5 PI, supplementation of HIGH up-regulated (P < 0.05) the mRNA expression of ZO1 and addition of CAR had greater (P < 0.05) mRNA expression of OCLN, compared with pigs in control group. On d 11 PI, supplementation of LOW or CAR had higher (P < 0.05) mRNA expression of CLDN1 in jejunal mucosa of weaned pigs, compared with the control and HIGH groups. On d 5 PI, supplementation of LOW down-regulated (P < 0.05) the mRNA expression of IL6, supplementation of HIGH had lower (P < 0.05) mRNA expression of IL1B, IL6, and TNF, and supplementation of CAR had lower (P < 0.05) IL1B and IL6 gene expression in ileal mucosa of weaned pigs in comparison to control pigs (Fig. 5). Supplementation of HIGH also had lower (P < 0.05) IL6 mRNA expression on d 11 PI in ileal mucosa, compared with the control group. However, no differences were observed in the gene expression of inflammatory mediators among LOW, HIGH, and CAR groups.

Gene expression profiles in jejunal mucosa of ETEC-infected weaned pigs fed diets supplemented with oligosaccharide-based polymer (Coligo) or antibiotics on d 5 or 11 post-inoculation (PI). a,bMeans without a common superscript differ (P < 0.05). Each least squares mean represents 6 observations. LOW = Low dose blood group A6 type 1-based polymer (Coligo); HIGH = High dose blood group A6 type 1-based polymer (Coligo); CAR = Carbadox; MUC2 = Mucin-2; CLDN1 = Claudin-1; ZO-1 = Zonula occludens-1; OCDN = Occludin

Gene expression profiles in ileal mucosa of ETEC-infected weaned pigs fed diets supplemented with oligosaccharide-based polymer (Coligo) or antibiotics on d 5 or 11 post-inoculation (PI). a,bMeans without a common superscript differ (P < 0.05). Each least squares mean represents 6 observations. LOW = Low dose blood group A6 type 1-based polymer (Coligo); HIGH = High dose blood group A6 type 1-based polymer (Coligo); CAR = Carbadox; IL1B: Interleukin-1 beta; IL6: Interleukin-6; TNF = Tumor necrosis factor; PTGS2: Cyclooxygenase-2
Discussion
ETEC infection is initiated by bacterial attachment to specific receptors on the intestinal epithelium by fimbrial adhesins, followed by colonization of ETEC in the small intestine [29]. Once colonization is established, ETEC rapidly proliferate and produce one or more enterotoxins, which can stimulate water and electrolyte secretion and reduce fluid absorption in the small intestine and induce diarrhea [30]. Diarrhea caused by ETEC is one of the most prevalent diseases during the weaning stage, which is responsible for anorexia, slower growth, or even the death of pigs. Results of the present study demonstrated that supplementation of Coligo improved growth rate, and reduced frequency of diarrhea and systemic inflammation of weaned pigs experimentally challenged with F18 ETEC. The potential mechanisms of action include inhibition of binding of bacteria and as such colonization of the gut by the F18 ETEC [11, 13], enhancing gut barrier function and reducing local and systemic inflammation.
In the current study, pigs in the control group grew slower and had a high frequency of diarrhea compared to pigs without ETEC challenge in our previous research [20, 56] and Priori et al. [57] suggest changes in the intestinal microbiota are affected by porcine blood group. Thus, the blood group A antigen in Coligo may affect the composition and function of microbial communities when fed to pigs. Moreover, recent studies demonstrated that dietary supplementation of ε-PL altered ileal microbiota structure and function in pigs [58] and fecal microbial community in mice [59]. ε-PL supplementation may promote the growth of beneficial microorganisms in the intestinal tract, therefore, reducing the proliferation of pathogens. The exact mechanisms of ε-PL in the current study remain unclear, so further research is needed to confirm the effects of Coligo on the pigs’ gut microbial community, intestinal inflammation, and immune responses against F18 ETEC. Pigs supplemented antibiotics also had reductions in mRNA expression of proinflammatory markers in the present study. This finding demonstrated that pigs supplemented antibiotics has less severe intestinal inflammation than pigs in control group. In agreement with previous research, antibiotic supplements might exert anti-inflammatory properties in the intestine or accumulate in phagocytic inflammatory cells, therefore, attenuating inflammatory responses in animals [60, 61]. Taken altogether, down-regulation in mRNA expression of proinflammatory cytokines by Coligo or antibiotics supplementation is beneficial for pigs in terms of their intestinal health and growth performance.
In conclusion, results in the current study suggest that in-feed supplementation of Coligo or antibiotic (carbadox) enhanced growth performance and reduced the severity of diarrhea caused by ETEC F18 infection. Although the percentage of β-hemolytic coliforms in fecal samples of pigs fed with Coligo was less diminished than pigs supplemented with antibiotics, enhanced disease resistance was demonstrated by the improved gut barrier integrity and attenuated systemic and intestinal inflammation. To further explore the mechanisms of action of Coligo, integrated metabolomics and metagenomics approaches may be considered to provide more insights into the beneficial effects of Coligo or other polymers on pigs’ health. Overall, the current study indicates that supplementation with Coligo has promising impacts on promoting growth and disease resistance of newly weaned pigs infected with ETEC F18. The efficacy of Coligo is comparable to antibiotic (carbadox) demonstrating the potential of Coligo as antibiotic alternative for animal growth performance and disease resistance. Large-scale animal trials are recommended to further evaluate the impacts of Coligo on performance of weaned pigs under commercial practice conditions.
Availability of data and materials
All data generated or analyzed during this study are available from the corresponding author upon reasonable request.
Abbreviations
- ADFI:
-
Average daily feed intake
- ADG:
-
Average daily gain
- BW:
-
Body weight
- CFU:
-
Colony-forming unit
- CLDN1 :
-
Claudin-1
- ETEC:
-
Enterotoxigenic E. coli
- IL1B :
-
Interleukin-1 beta
- IL6 :
-
Interleukin-6
- mRNA:
-
Messenger RNA
- MUC2 :
-
Mucin-2
- OCDN :
-
Occludin
- PI:
-
Post-inoculation
- qRT-PCR:
-
Quantitative real-time polymerase chain reaction
- TNF :
-
Tumor necrosis factor
- ZO-1 :
-
Zona occludens-1
References
Nagy B, Fekete PZ. Enterotoxigenic Escherichia coli in veterinary medicine. Int J Med Microbiol. 2005;295(6-7):443–54. https://doi.org/10.1016/j.ijmm.2005.07.003.
Fairbrother JM, Nadeau É, Gyles CL. Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim Health Res Rev. 2005;6(1):17–39. https://doi.org/10.1079/AHR2005105.
Van Boeckel TP, Brower C, Gilbert M, Grenfell BT, Levin SA, Robinson TP, et al. Global trends in antimicrobial use in food animals. Proc Natl Acad Sci. 2015;112(18):5649–54. https://doi.org/10.1073/pnas.1503141112.
FDA (Food and Drug Administration) New animal drugs and new animal drug combination products administered in or on medicated feed or drinking water of food-producing animals: recommendations for drug sponsors for voluntarily aligning product use conditions with FDA Guidance for Industry #213. Center for Veterinary Medicine. Washington, DC: US Department of Health and Human Services; 2016. https://www.fda.gov/downloads/AnimalVeterinary/GuidanceComplianceEnforcement/GuidanceforIndustry/UCM299624.pdf.
Nollet H, Deprez P, Van Driessche E, Muylle E. Protection of just weaned pigs against infection with F18+ Escherichia coli by non-immune plasma powder. Vet Microbiol. 1999;65(1):37–45. https://doi.org/10.1016/S0378-1135(98)00282-X.
Newburg DS. Do the binding properties of oligosaccharides in milk protect human infants from gastrointestinal bacteria. J Nutr. 1997;123:980S–4S. https://doi.org/10.1093/jn/127.5.980S.
Coppa GV, Zampini L, Galeazzi T, Facinelli B, Ferrante L, Capretti R, et al. Human milk oligosaccharides inhibit the adhesion to Caco-2 cells of diarrheal pathogens: Escherichia coli, vibrio cholerae, and Salmonella fyris. Pediatr Res. 2006;59(3):377–82. https://doi.org/10.1203/01.pdr.0000200805.45593.17.
Lindahl M, Wadström T. K99 surface haemagglutinin of enterotoxigenic E. coli recognize terminal n-acetylgalactosamine and sialic acid residues of glycophorin and other complex glycoconjugates. Vet Microbiol. 1984;9(3):249–57. https://doi.org/10.1016/0378-1135(84)90042-7.
Erickson AK, Baker DR, Bosworth BT, Casey TA, Benfield DA, Francis DH, et al. Characterization of porcine intestinal receptors for the K88ac fimbrial adhesin of Escherichia coli as mucin-type sialoglycoproteinst K88ac+ enterotoxigenic Escherichia coli infections. Infect Immun. 1994;62(12):5404–10. https://doi.org/10.1128/iai.62.12.5404-5410.1994.
Coddens A, Verdonck F, Tiels P, Rasschaert K, Goddeeris BM, Cox E. The age-dependent expression of the F18+ E. coli receptor on porcine gut epithelial cells is positively correlated with the presence of histo-blood group antigens. Vet Microbiol. 2007;122(3-4):332–41. https://doi.org/10.1016/j.vetmic.2007.02.007.
Coddens A, Diswall M, Ångström J, Breimer ME, Goddeeris B, Cox E, et al. Recognition of blood group ABH type 1 determinants by the FedF adhesin of F18-fimbriated Escherichia coli. J Biol Chem. 2009;284(15):9713–26. https://doi.org/10.1074/jbc.M807866200.
Moonens K, Bouckaert J, Coddens A, Tran T, Panjikar S, De Kerpel M, et al. Structural insight in histo-blood group binding by the F18 fimbrial adhesin FedF. Mol Microbiol. 2012;86(1):82–95. https://doi.org/10.1111/j.1365-2958.2012.08174.x.
Coddens A, Cox E, Teneberg SE. Inhibitors of f18+ E. coli binding. European patent office. European patent no. EP2344167B1. 2014. https://worldwide.espacenet.com/patent/search?q=pn%3DEP2344167B1
Lin K, Kasko AM. Carbohydrate-based polymers for immune modulation. ACS Macro Lett. 2014;3(7):652–7. https://doi.org/10.1021/mz5002417.
Ekladious I, Colson YL, Grinstaff MW. Polymer–drug conjugate therapeutics: advances, insights and prospects. Nat Rev Drug Discov. 2019;18(4):273–94. https://doi.org/10.1038/s41573-018-0005-0.
Shukla SC, Singh A, Pandey AK, Mishra A. Review on production and medical applications of ɛ-polylysine. Biochem Eng J. 2012;65:70–81. https://doi.org/10.1016/j.bej.2012.04.001.
Yuan J, Guo L, Wang S, Liu D, Qin X, Zheng L, et al. Preparation of self-assembled nanoparticles of ε-polylysine-sodium alginate: a sustained-release carrier for antigen delivery. Colloids Surf B Biointerfaces. 2018;171:406–12. https://doi.org/10.1016/j.colsurfb.2018.07.058.
Kreuzer S, Reissmann M, Brockmann GA. New fast and cost-effective gene test to get the ETEC F18 receptor status in pigs. Vet Microbiol. 2013;163(3-4):392–4. https://doi.org/10.1016/j.colsurfb.2018.07.058.
National Research Council (NRC). Nutrient requirements of swine: 11th revised edition. Washington, DC: The National Academies Press; 2012. https://doi.org/10.17226/13298.
Liu Y, Song M, Che TM, Almeida JAS, Lee JJ, Bravo D, et al. Dietary plant extracts alleviate diarrhea and alter immune responses of weaned pigs experimentally infected with a pathogenic Escherichia coli. J Anim Sci. 2013;91(11):5294–306. https://doi.org/10.2527/jas.2012-6194.
Kim K, Ehrlich A, Perng V, Chase JA, Raybould H, Li X, et al. Algae-derived β-glucan enhanced gut health and immune responses of weaned pigs experimentally infected with a pathogenic E. coli. Anim Feed Sci Technol. 2019;248:114–25. https://doi.org/10.1016/j.anifeedsci.2018.12.004.
Kim K, He Y, **ong X, Ehrlich A, Li X, Raybould H, et al. Dietary supplementation of Bacillus subtilis influenced intestinal health of weaned pigs experimentally infected with a pathogenic E. coli. J Anim Sci Biotechnol. 2019;10:52. https://doi.org/10.1186/s40104-019-0364-3.
Almeida JAS, Liu Y, Song M, Lee JJ, Gaskins HR, Maddox CW, et al. Escherichia coli challenge and one type of smectite alter intestinal barrier of pigs. J Anim Sci Biotechnol. 2013;4:52. https://doi.org/10.1186/2049-1891-4-52.
Garas LC, Feltrin C, Kristina Hamilton M, Hagey JV, Murray JD, Bertolini LR, et al. Milk with and without lactoferrin can influence intestinal damage in a pig model of malnutrition. Food Funct. 2016;7(2):665–78. https://doi.org/10.1039/c5fo01217a.
DebRoy C, Maddox CW. Identification of virulence attributes of gastrointestinal Escherichia coli isolates of veterinary significance. Anim Health Res Rev. 2001;2(2):129–40. https://doi.org/10.1079/AHRR200131.
Deplancke B, Gaskins HR. Microbial modulation of innate defense: goblet cells and the intestinal mucus layer. Am J Clin Nutr. 2001;73(6):1131–41. https://doi.org/10.1093/ajcn/73.6.1131S.
Liu Y, Song M, Che TM, Lee JJ, Bravo D, Maddox CW, et al. Dietary plant extracts modulate gene expression profiles in ileal mucosa of weaned pigs after an Escherichia coli infection. J Anim Sci. 2014;92(5):2050–62. https://doi.org/10.2527/jas.2013-6422.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25(4):402–8. https://doi.org/10.1006/meth.2001.1262.
Sellwood R, Gibbons RA, Jones GW, Rutter JM. Adhesion of enteropathogenic Escherichia coli to pig intestinal brush borders: the existence of two pig phenotypes. J Med Microbiol. 1975;8(3):405–11. https://doi.org/10.1099/00222615-8-3-405.
Nagy B, Zs FP. Enterotoxigenic Escherichia coli (ETEC) in farm animals. Vet Res. 1999;30(2-3):259–84. Available from: https://pubmed.ncbi.nlm.nih.gov/10367358.
Cheng G, Sa W, Cao C, Guo L, Hao H, Liu Z, et al. Quinoxaline 1,4-di-N-oxides: biological activities and mechanisms of actions. Front Pharmacol. 2016;7:64–85. https://doi.org/10.3389/fphar.2016.00064.
Das NK. In vitro susceptibility of Escherichia coli of swine origin to carbadox and other antimicrobials. Am J Vet Res. 1984;45(2):252–4. Available from: https://pubmed.ncbi.nlm.nih.gov/6370050.
Holmner A, Askarieh G, Okvist M, Krengel U. Blood group antigen recognition by Escherichia coli heat-labile enterotoxin. J Mol Biol. 2007;371(3):754–64. https://doi.org/10.1016/j.jmb.2007.05.064.
Barra JL, Monferran CG, Balanzino LE, Cumar FA. Escherichia coli heat-labile enterotoxin preferentially interacts with blood group A-active glycolipids from pig intestinal mucosa and A- and B-active glycolipids from human red cells compared to H-active glycolipids. Mol Cell Biochem. 1992;115(1):63–70. https://doi.org/10.1007/BF00229097.
Lee SH. Intestinal permeability regulation by tight junction: implication on inflammatory bowel diseases. Intest Res. 2015;13(1):11–8. https://doi.org/10.5217/ir.2015.13.1.11.
Berkes J, Viswanathan VK, Savkovic SD, Hecht G. Intestinal epithelial responses to enteric pathogens: effects on the tight junction barrier, ion transport, and inflammation. Gut. 2003;52(3):439–51. https://doi.org/10.1136/gut.52.3.439.
Dubreuil JD. Enterotoxigenic Escherichia coli targeting intestinal epithelial tight junctions: an effective way to alter the barrier integrity. Microb Pathog. 2017;113:129–34. https://doi.org/10.1016/j.micpath.2017.10.037.
Rose R, Whipp SC, Moon HW. Effects of Escherichia coli heat-stable enterotoxin b on small intestinal villi in pigs, rabbits, and lambs. Vet Pathol. 1987;24(1):71–9. https://doi.org/10.1177/030098588702400112.
Balda MS, Matter K. Tight junctions at a glance. J Cell Sci. 2008;121(22):3677–82. https://doi.org/10.1242/jcs.023887.
Nagpal R, Yadav H. Bacterial translocation from the gut to the distant organs: an overview. Ann Nutr Metab. 2017;71(Suppl. 1):11–6. https://doi.org/10.1159/000479918.
Berg RD. Bacterial translocation from the gastrointestinal tract. Trends Microbiol. 1995;3(4):149–54. https://doi.org/10.1016/S0966-842X(00)88906-4.
Berg RD. Bacterial translocation from the gastrointestinal tract. J Med. 1992;23(3-4):217–44. Available from: https://pubmed.ncbi.nlm.nih.gov/1479301.
Lessard M, Dupuis M, Gagnon N, Nadeau E, Matte JJ, Goulet J, et al. Administration of Pediococcus acidilactici or Saccharomyces cerevisiae boulardii modulates development of porcine mucosal immunity and reduces intestinal bacterial translocation after Escherichia coli challenge. J Anim Sci. 2009;87(3):922–34. https://doi.org/10.2527/jas.2008-0919.
He Y, **no C, Kim K, Wu Z, Tan B, Li X, et al. Dietary Bacillus spp. enhanced growth and disease resistance of weaned pigs by modulating intestinal microbiota and systemic immunity. J Anim Sci Biotechnol. 2020;11(1):101–20. https://doi.org/10.1186/s40104-020-00498-3.
Dubreuil JD, Isaacson RE, Schifferli DM. Animal enterotoxigenic Escherichia coli. EcoSal Plus. 2016;7:1–80. https://doi.org/10.1128/ecosalplus.ESP-0006-2016.
Bannerman DD, Goldblum SE. Mechanisms of bacterial lipopolysaccharide-induced endothelial apoptosis. Am J Physiol Lung Cell Mol Physiol. 2003;284(6):L899–914. https://doi.org/10.1152/ajplung.00338.2002.
Song M, Liu Y, Soares JA, Che TM, Osuna O, Maddox CW, et al. Dietary clays alleviate diarrhea of weaned pigs. J Anim Sci. 2012;90(1):345–60. https://doi.org/10.2527/jas.2010-3662.
Medzhitov R, Janeway CA Jr. Innate immunity: impact on the adaptive immune response. Curr Opin Immunol. 1997;9(1):4–9. https://doi.org/10.1016/S0952-7915(97)80152-5.
Sansonetti PJ. War and peace at mucosal surfaces. Nat Rev Immunol. 2004;4(12):953–64. https://doi.org/10.1038/nri1499.
Pié S, Lallès JP, Blazy F, Laffitte J, Sève B, Oswald IP. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J Nutr. 2004;134(3):641–7. https://doi.org/10.1093/jn/134.3.641.
Dinarello CA. Proinflammatory cytokines. Chest. 2000;118(2):503–8. https://doi.org/10.1378/chest.118.2.503.
Devriendt B, Stuyven E, Verdonck F, Goddeeris BM, Cox E. Enterotoxigenic Escherichia coli (K88) induce proinflammatory responses in porcine intestinal epithelial cells. Dev Comp Immunol. 2010;34(11):1175–82. https://doi.org/10.1016/j.dci.2010.06.009.
Lodemann U, Amasheh S, Radloff J, Kern M, Bethe A, Wieler LH, et al. Effects of ex vivo infection with ETEC on jejunal barrier properties and cytokine expression in probiotic-supplemented pigs. Dig Dis Sci. 2017;62(4):922–33. https://doi.org/10.1007/s10620-016-4413-x.
Zanello G, Meurens F, Berri M, Chevaleyre C, Melo S, Auclair E, et al. Saccharomyces cerevisiae decreases inflammatory responses induced by F4+ enterotoxigenic Escherichia coli in porcine intestinal epithelial cells. Vet Immunol Immunopathol. 2011;141(1-2):133–8. https://doi.org/10.1016/j.vetimm.2011.01.018.
McLamb BL, Gibson AJ, Overman EL, Stahl C, Moeser AJ. Early weaning stress in peigs impairs innate mucosal immune responses to enterotoxigenic E coli challenge and exacerbates intestinal injury and clinical disease. PLoS One. 2013;8(4):e59838. https://doi.org/10.1371/journal.pone.0059838.
Motta V, Luise D, Bosi P, Trevisi P. Faecal microbiota shift during weaning transition in piglets and evaluation of AO blood types as sha** factor for the bacterial community profile. PLoS One. 2019;14(5):e0217001. https://doi.org/10.1371/journal.pone.0217001.
Priori D, Colombo M, Koopmans S-J, Jansman AJM, van der Meulen J, Trevisi P, et al. The AO blood group genotype modifies the jejunal glycomic binding pattern profile of piglets early associated with a simple or complex microbiota. J Anim Sci. 2016;94(2):592–601. https://doi.org/10.2527/jas.2015-9948.
Zhang X, Hou Z, Xu B, **e C, Wang Z, Yu X, et al. Dietary supplementation of ε-polylysine beneficially affects ileal microbiota structure and function in Ningxiang pigs. Front Microbiol. 2020;11:2940. https://doi.org/10.3389/fmicb.2020.544097.
You X, Einson JE, Lopez-Pena CL, Song M, **ao H, McClements DJ, et al. Food-grade cationic antimicrobial ε-polylysine transiently alters the gut microbial community and predicted metagenome function in CD-1 mice. NPJ Sci Food. 2017;1(1):8–18. https://doi.org/10.1038/s41538-017-0006-0.
Niewold TA. The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poult Sci. 2007;86(4):605–9. https://doi.org/10.1093/ps/86.4.605.
Costa E, Uwiera RRE, Kastelic JP, Selinger LB, Inglis GD. Non-therapeutic administration of a model antimicrobial growth promoter modulates intestinal immune responses. Gut Pathog. 2011;3(1):14–29. https://doi.org/10.1186/1757-4749-3-14.
Acknowledgments
Not applicable.
Funding
This project was supported by Pancosma SA, Geneva, Switzerland and the United States Department of Agriculture (USDA) National Institute of Food and Agriculture (NIFA), multistate projects W4002 and NC1202.
Author information
Authors and Affiliations
Contributions
The contributions of the authors were as follows: KK conducted the experiment and wrote the manuscript. YH, CJ, and LK, and AE helped to conduct animal trial and part of the laboratory work and helped to revise the manuscript. XL provided ETEC inoculum and helped to revise the manuscript. DB and EC revised the manuscript. YL was the principal investigator. YL designed the experiment, oversaw the development of the study and wrote the last version of the manuscript. The authors declare no conflicts of interest. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The protocol for this study was reviewed and approved by the Institutional Animal Care and Use Committee at the University of California, Davis (IACAC #19322). The study was conducted at the Cole Facility at the University of California, Davis.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Supplementary Information
Additional file 1.
Table S1 Gene-specific primer sequences and PCR conditions.
Additional file 2.
Table S2 Red blood cell profiles of ETEC-infected weaned pigs fed diets supplemented with oligosaccharide-based polymer (Coligo) or antibiotics.
Additional file 3.
Table S3 Intestinal morphology of ETEC-infected weaned pigs fed diets supplemented with oligosaccharide-based polymer (Coligo) or antibiotics.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kim, K., He, Y., **no, C. et al. Supplementation of oligosaccharide-based polymer enhanced growth and disease resistance of weaned pigs by modulating intestinal integrity and systemic immunity. J Animal Sci Biotechnol 13, 10 (2022). https://doi.org/10.1186/s40104-021-00655-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-021-00655-2