
Abstract
Background
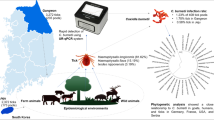
Rickettsia spp. are important tick-borne pathogens that cause various human and animal diseases worldwide. A tool for rapid and accurate detection of the pathogens from its vectors is necessary for prevention of Rickettsioses propagation in humans and animals, which are infested by ticks. Therefore, this study was conducted to evaluate a molecular tool, ultra-rapid real-time PCR (UR-qPCR), for rapid and accurate detection of Rickettsia spp. from 5644 ticks in 408 pools collected from livestock and their surrounding environments in Gangwon and Jeju province in South Korea.
Results
The UR-qPCR of Rickettsia DNA showed a limit of detection of 2.72 × 101 copies of Rickettsia DNA and no cross reaction with other tick-borne pathogens, namely Anaplasma phagocytophilum, Ehrlichia chaffeensis, E. canis, Toxoplasma gondii, and Borrelia burgdorferi. In addition, the PCR assay also showed possibility of various Rickettsia species detection including R. monacensis, “Candidatus R. longicornii”, R. japonica, R. roultii, and R. tamurae. The collected ticks were identified with major species belonged to Haemaphysalis longicornis (81.62%), followed by H. flava (15.19%), and Ixodes nipponensis (3.19%). Rickettsia detection from tick samples using the UR-qPCR showed that the minimum infection rate (MIR) of Rickettsia in collected ticks was 1.24‰ and that all positive pools contained H. longicornis, equal to the MIR of 1.39‰ of this species. Additionally, MIR of Rickettsia spp. detected in ticks collected in Gangwon and Jeju was 1.53‰ and 0.84‰, respectively. Furthermore, the sequencing results of the 17 kDa protein antigen gene and ompA gene showed that Rickettsia spp. sequences from all pools were related to “Candidatus R. longicornii” and “Candidatus R. **gxinensis”.
Conclusions
The UR-qPCR system was demonstrated to be useful tool for accurate and rapid detection of Rickettsia from its vector, ixodid ticks, within 20 min. The data on Rickettsia spp. in ticks detected in this study provide useful information on the distribution of Rickettsia in previously unstudied Korean provinces, which are important for the prevention and control of the spread of rickettsioses in both animals and humans in the country.
Similar content being viewed by others


Background
The obligate intracellular bacteria of Rickettsia genus are commonly harboured and transmitted by arthropods, mainly ticks [1,2,3], some of the bacteria cause Rickettsioses in animals and humans with mild to life-threatening consequences [4]. Rickettsia and the related tick vectors have been reported in different countries. For example, four subspecies of R. conorii, the cause Mediterranean spotted fever in Europe, were mainly found in Rhipicephalus sanguineus and Rh. pumilio ticks [5, 6], the Japan spotted fever group (R. tamurae, R. japonica, R. raoultii, and Candidatus R. principis) was detected in Haemaphysalis and Amblyomma ticks in Japan [7], and R. raoultii was prevalent in Dermacentor nuttalli and Dermacentor silvarum ticks in China [8, 9]. The information of Rickettsia species and related tick species in a particular region is important to identify the risk of Rickettsioses transmission via tick bite.
Polymerase chain reaction (PCR) has been used as a sensitive and specific tool for the rapid detection of Rickettsia from both ticks and patients [10,11,12,13,14], and species identification of Rickettsia was done by sequencing analysis of various genes, such as the rickettsial citrate synthase gene (gltA) [15], SFGR-specific 190 kDa outer membrane protein A gene (ompA) [16], outer-membrane protein rOmpB (OmpB) [17], surface cell antigen gene “gene D” (Sca4) [18, 19], and the genus-specific 17 kDa outer membrane antigen gene [20]. Afterwards, real-time PCR was demonstrated to be more sensitive and rapid compared to conventional nested PCR for Rickettsia detection, and become an important tool for screening of Rickettsia from its natural reservoirs or vectors [21,22,23]. However, the current Rickettsia detection real-time PCRs are still time-consuming systems. A new chip-based PCR system named ultra-rapid real-time PCR (UR-qPCR) has been developed, which has optimal thermal transfer with chip-based reaction that reduces turnaround time. In addition, this small-footprint device with low power consumption make it possible for point-of-care testing application. It has been shown to be useful for the sensitive and rapid detection of honeybee pathogens on-site [24,25,26]. Therefore, the UR-qPCR could be a useful tool for rapid detection of Rickettsia from ticks.
In Korea, Rickettsioses in humans have been reported since 2006 [27, 28]. Thereafter, the role of ticks in carrying and transmission of Rickettsia spp. to humans was also demonstrated [29,30,31,32,33,34]. Monitoring of Rickettsia in ticks was done in northern and western regions of the country and showed that “Candidatus R. longicornii” was the most prevalent Rickettsia species carried by ticks [30, 35, 36]. However, the information of Rickettsia harboured by ticks in other regions of the country is still remained uncharacterized.
Accordingly, this study was conducted to examine the ability of the UR-qPCR system for detection of Rickettsia in ticks collected from wild animals and livestock in two provinces: Gangwon and Jeju, located in northeastern and southern region of South Korea, respectively. Sequencing and phylogenetic analyses of detected Rickettsia spp. were done using the 17 kDa protein antigen and ompA genes.
Results
Sensitivity and specificity of Rickettsia UR-qPCR
Amplification using serially diluted recombinant DNA showed a limit of detection of 2.72 × 101 copies of Rickettsia DNA (Fig. 1A and B). The linear regression representing the relationship between initial DNA copy and cycle threshold (Ct) of amplification was established from triplicate PCR reactions, y = − 3.5171x + 42.424; R2 = 0.9966, where x and y are the log10 DNA copy number and Ct value, respectively (Fig. 1C). The amplification efficiency calculated from the slope of the standard curve (E = 10(− 1/slope)-1) was 92.45%. Furthermore, the peaks of melting temperature of amplification (Fig. 2A) showed that the UR-qPCR can be used for specific detection of Rickettsia among the tested DNA templates originating from other tick-borne pathogens, namely Anaplasma phagocytophilum, Ehrlichia chaffeensis, E. canis, Toxoplasma gondii, Coxiella burnetii, and Borrelia burgdorferi. In addition, the melting peaks of five different Rickettsia spp. (R. japonica, R. roultii, “Candidatus R. longicornii”, R. monacensis, and R. tamurae) detection were not greatly different, ranging from 76.03 °C to 77.01 °C (Fig. 2B). The result demonstrated that the UR-qPCR assay can be used as a molecular tool for detection of various Rickettsia species.

Sensitivity of detection of Rickettsia species using ultra-rapid real-time PCR (UR-qPCR). Amplification curves of UR-qPCR using 2.72 × 108 to 2.72 × 100 copies of Rickettsia DNA (denoted by number 8 to 0; A). The melting curves show Rickettsia detection is possible from 2.72 × 108 to 2.72 × 101 copies of target DNA (number 8–1; B). “N” is the negative control without a DNA template. Linear regression representing the relationship between cycle threshold of amplification (Ct value) and initial DNA copy number (C) was established by amplifying 10-fold dilutions of Rickettsia DNA from 2.72 × 108 to 2.72 × 101 DNA copies in triplicate

Specificity of detection of Rickettsia species using ultra-rapid real-time PCR (UR-qPCR). The specificity of Rickettsia UR-qPCR is demonstrated by different melting temperatures observed when amplifying Rickettsia japonica recombinant DNA, Rickettsia sp. DNA from total nucleic acids isolated from tick sample, and the DNA of other common tick-borne pathogens, namely Anaplasma phagocytophilum, Ehrlichia chaffeensis, E. canis, Toxoplasma gondii, Borrelia burgdorferi, Coxiella burnetii, and samples with no DNA template (A). The melting temperature ranging from 76.03 °C to 77.01 °C were seen from detection PCR using DNA template of five different Rickettsia species (B). “N1” and “N2” are negative result using total nucleic acids isolated from two tick pools, and “N” is negative control without DNA template
Prevalence of Rickettsia species in ticks
The tick species identified from the highest number of pools were Haemaphysalis longicornis (333/408 pools; 81.62%), followed by H. flava (62/408 pools; 15.19%), and Ixodes nipponensis (13/408 pools; 3.19%); H. longicornis and H. flava were present in samples collected from both Gangwon and Jeju provinces, whereas I. nipponensis was only detected in samples collected from Gangwon province.
Moreover, among the three most common tick species identified, only H. longicornis from both provinces harboured Rickettsia spp. The minimum infection rate (MIR) in Gangwon province as determined by ITS DNA detection using UR-qPCR and by 17 kDa protein antigen and ompA gene detection using conventional nested PCR was 1.53‰ (5/408 pools), 1.22‰ (4/408 pools), and 1.53‰ (5/408 pools), respectively. In Jeju province, the MIR was 0.84‰ (2/408 pools), 2.53‰ (6/408 pools), and 2.53‰ (6/408 pools) as detected by ITS, 17 kDa protein antigen gene, and ompA detection, respectively (Table 1).
The overall MIR was 1.24‰ (7/408 pools), 1.77‰ (10/408 pools), and 1.95‰ (11/408 pools) for the detection methods targeting ITS, 17 kDa protein antigen gene, and ompA gene, respectively (Table 1). The MIR according to the developmental stages of the infected tick species ranged from 4.81–9.62‰ for nymphs, 21.89‰ for adult males, and 6.70–13.39‰ for adult females; the prevalence was 0% for larvae.
Sequencing and phylogenetic analysis
Among the 10 pools, generated sequences of the 17 kDa protein antigen gene were 100.00% identical to each other (NCBI accession No.: MW916824) and had 100.00% identity with NCBI deposited sequences of “Candidatus R. longicornii” and “Candidatus R. **gxinensis” that had been detected in H. longicornis ticks in Korea and China, respectively. Additionally, the sequences of the ompA gene (NCBI accession No.: MW916823) were 100.00% identical among all 11 pools and showed 100.00% identity to sequences of “Candidatus R. longicornii” and “Candidatus R. **gxinensis” detected from H. longicornis ticks in Korea and China, respectively. Phylogenetic analysis of the two genes showed that the detected Rickettsia spp. clustered with “Candidatus R. **gxinensis” and “Candidatus R. longicornii” when compared to Rickettsia spp. originating from other countries (Fig. 3).

Phylogenetic trees of Rickettsia species. The trees were created based on the 410 bp 17 kDa protein antigen gene (A) and the 488 bp ompA gene (B) of Rickettsia species using the neighbour-joining method and bootstrap analysis (1000 reiterations) carried out according to the Kimura 2-parameter method in MEGA7 software
Discussion
Screening of Rickettsia was done from ticks collected from Gangwon province, located in northeastern Korea, and from Jeju island, located in southern Korea. Only H. longicornis ticks were found to be the vector of Rickettsia with positive result of 11/408 pools detected by ompA gene. The detected Rickettsia spp. all showed 100% sequence similarity to “Candidatus R. longicornii” and “Candidatus R. **gxinensis”. The result of this study was consistent with previous report [35] that only “Candidatus R. longicornii” was detected in H. longicornis in the northern area near the demilitarized zone in South Korea. However, various Rickettsia species were detected in H. longicornis ticks in other regions of the country, such as R. japonica, R. heilongjiangensis, and R. monacensis in northwestern and southwestern provinces [30], “Candidatus R. longicornii” and R. koreansis in midwestern region [36]. Rickettsia spp. were also minorly detected in H. flava and I. nipponensis in these regions. The result of Rickettsia detection in this study could be important to fulfil the knowledge of distribution of Rickettsia and its vectors in the country.
“Candidatus R. **gxinensis” and “Candidatus R. longicornii” were detected mainly in China and South Korea, in which the name “Candidatus R. **gxinensis” was first proposed as a potential new species based on the analysis of gltA and ompA genes [8] for a Rickettsia sp. detected in H. longicornis ticks collected at **gxin town in **lin province, China. This species was then identified in Rhipicephalus microplus tick in China and in H. longicornis in South Korea [37, 38]. The other proposed species, “Candidatus R. longicornii”, was introduced based on the analysis of rrs, gltA, ompA, ompB, and sca4 genes [35]. This species was also prevalently detected in H. longicornis tick distributed in South Korea, and China [38,39,37]. Therefore, “Candidatus R. **gxinensis” and “Candidatus R. longicornii” could be the only one species, and according to the prevalence of the organisms detected in H. longicornis tick, the only name “Candidatus R. longicornii” should be used for the Rickettsia species.
The distribution of tick species identified from Gangwon and Jeju provinces was the same as that reported in other provinces; H. longicornis was the most abundant species among the three most common tick species in Korea - H. longicornis, H. flava, and I. nipponensis [41, 42]. The H. longicornis tick is a common parasite of livestock, wild animals, and humans, and it is distributed in ten countries including eastern Asia, the USA, Australia, and New Zealand [35, 43]. H. longicornis ticks were known to be vectors of various diseases including rickettsioses [43,44,45].
The loop-mediated isothermal amplification (LAMP) assay was developed for rapid detection of Rickettsia from ticks within 30 min [46], using the crude DNA prepared by heating method [47] the LAMP was demonstrated to useful for on-site detection of Rickettsia from vectors. However, using hydroxynaphthol blue as a colorimetric component for visual detection by naked eye in LAMP could make a challenge for different readers, and therefore the results need to be confirmed in electrophoresis [46]. The UR-qPCR evaluated in this study showed possibility of various Rickettsia species detection within 20 min could address the disadvantages of LAMP for accurate and rapid detection of Rickettsia. Using the crude preparation of DNA [47] the UR-qPCR could be used for on-site screening of Rickettsia from ticks.
Conclusions
In this study, a molecular tool UR-qPCR for the rapid detection of Rickettsia spp. in ticks was initially examined. The PCR system showed a limit detection of around 27.2 copies of Rickettsia DNA within around 20 min. The possibility of various Rickettsia species detection was confirmed, and the usefulness of Rickettsia spp. detection was also demonstrated from tick samples. The rapidity and mobility of this PCR system could be important to develop a molecular tool for on-site detection of Rickettsia sp. from its vectors. Additionally, the prevalence data on Rickettsia spp. identified in ticks collected from livestock and wild animals in the Gangwon and Jeju provinces provide useful information on Rickettsia distribution in previously unstudied Korean provinces; this is important for the prevention and control of the spread of rickettsioses in both animals and humans in the country.
Methods
Tick sample collection
A total of 5644 larval, nymphal, and adult ticks were collected from livestock, wild animals, and vegetation surrounding the farms or living areas of wild animal in the Gangwon and Jeju provinces in Korea between August and November in 2019. Species of ticks were identified by their morphological characteristics using a stereomicroscope (Discovery.V8; ZEISS, Oberkochen, Germany) and the standard illustrated taxonomic key [48]. After identification of species, the samples were pooled for the living stages of the same species collected from the same site. Each pool contained 1, 1 to 10, and 1 to 50 individuals of adult, nymph, and larvae, respectively. Totally, 408 pools were acquired, in which 235 pools were collected from Gangwon and designated as 19 M1 to 19 M235, while 173 pools were collected from Jeju and labelled as 19 T1 to 19 T173. The samples were then preserved in 70% ethanol and stored at − 80 °C until further analysis.
Nucleic acid extraction from ticks
Briefly, ticks from each pool were washed three times using the UltraPure™ distilled water (Thermo Fisher Scientific, Massachusetts, USA), and were placed in a tissue homogeniser with steel beads 2.381 mm diameter (SNC, Hanam, Korea). After adding 600 μl of PBS solution, the sample was homogenised using a Precellys 24 Tissue Homogeniser (Bertin Instruments, Montigny-le-Bretonneux, France). Then 300 μl of the homogenate was transferred to a new tube that contained 300 μl of lysis buffer and 30 μl of proteinase K solution. The mixture was incubated at 56 °C for 10 min and the total nucleic acid was extracted using the Maxwell® RSC Viral Total Nucleic Acid Purification Kit on the automated Maxwell® RSC Instrument (Promega, Madison, WI, USA) according to the manufacturer’s instructions. Finally, 50 μl of total nucleic acid was acquired from each sample.
PCR performance
The internal transcribed spacer (ITS) region of Rickettsia spp. was targeted for detection in tick samples using the GENECHECKER® UF-150 UR-qPCR system (Genesystem Co., Ltd., Daejeon, Korea) and 2× Rapi: Detect™ Master mix with dye (SYBR green, Cat. No.: 9799100100; Genesystem Co.). The 10 μl reaction mix consisted of 1 μl (10 pmol) of each primer (ITS-F/R; Table 2), 5 μl of PCR premix, and 3 μl of total nucleic acid. The PCR conditions were examined at different annealing temperature from 52 °C to 66 °C to select the optimal condition for specific and sensitive detection of Rickettsia spp., final PCR conditions are shown in Table 2. Recombinant ITS DNA of R. japonica (NCBI accession number CP047359) was used for optimizing PCR conditions and was used as positive control for Rickettsia spp. detection from tick samples, and no DNA template was used in negative control.
Species identification was performed by nested PCR using the Mastercycler® X50s conventional PCR system (Eppendorf, Hamburg, Germany) and sequence analysis of the ompA gene [50, 51] and the 17 kDa protein antigen gene [50] using the AccuPower® ProFi Taq PCR Premix (Bioneer, Daejeon, Korea). The 20 μl reaction mix was composed of 1 μl (10 pmol) of each primer (Table 2), 13 μl of ddH2O, and 5 μl of total nucleic acid. The PCR conditions are shown in Table 2. No DNA template was used in negative control.
The sensitivity of rickettsial UR-qPCR was examined using the recombinant DNA of R. japonica, a DNA fragment 388 bp long corresponding to the amplicon size of primer pair ITS-F/R was chemically synthesized according to the sequence from position 700,066 to 700,453 of R. japonica genome (NCBI accession number CP047359). The DNA fragment consisting of 92 bp of 23S ribosomal RNA gene, 253 bp internal transcribed spacer, 43 bp of 5S ribosomal RNA gene was inserted in the pGEM®-T vector system (Promega, Madison, WI, USA) and used as standard DNA for positive control of UR-qPCR detection. Recombinant DNA was serially 10-fold diluted from 2.72 × 108 to 2.72 × 100 copies/μl, and used for UR-qPCR to identify the minimum copy number that could be detected. PCR was performed in triplicate and a linear regression representing the relationship between initial DNA copy number and cycle threshold (Ct) of amplification was established. The specificity of the UR-qPCR system was also evaluated by assessing cross-detection of DNA from five other tick-borne pathogens (Anaplasma phagocytophilum, Ehrlichia chaffeensis, E. canis, Toxoplasma gondii, Coxiella burnetii, and Borrelia burgdorferi) under the same PCR conditions. Specific amplification was determined by comparing the peak of melting curves using sample DNA with that using Rickettsia recombinant DNA. The UR-qPCR assay was also performed using DNA template of five Rickettsia species including R. japonica, R. roultii, “Candidatus R. longicornii”, R. monacensis, and R. tamurae to verify the possibility of various Rickettsia species detection.
Detection and phylogenetic analysis of Rickettsia from ticks
To screen ticks for Rickettsia spp., 10 μl total nucleic acid from each of the five tick pools of adults, nymphs, or larvae of the same species collected from the same site was taken and combined to have 50 μl solution mix; then, 3 μl was used for UR-qPCR. The combined nucleic acid with positive UR-qPCR results was identified and each pool tested individually to identify the exact pool carrying the pathogen; conventional nested PCRs targeting ompA and 17 kDa protein antigen genes were used for the detection and sequencing analysis.
After confirming the expected bands of ITS DNA (388 bp) and nested PCR products of ompA (532 bp, Table 2) and 17 kDa protein antigen genes (450 bp, Table 2) the PCR products were purified using the QIAquick PCR Purification Kit (QIAGEN, Hilden, Germany) prior to being shipped for sequencing by Macrogen Inc. (Seoul, Korea). The generated sequences were deposited on NCBI with accession number MW916824 (17 kDa protein antigen gene), MW916823 (ompA gene), and MW929192 (ITS DNA). The gene sequences were aligned using the Clustal X2 program [52], the overhanging ends were trimmed using BioEdit 7.2 software [53], and phylogenetic tree was constructed using the neighbour-joining method and bootstrapped 1000 times using the MEGA7 software [54].
Statistical analysis
The tick samples were collected and arranged in pools according to living stages of each species collected from the same site for detection of Rickettsia spp. Analysis of the prevalence of Rickettsia spp. in the tick pools was done using the minimum infection rate (MIR) that based on the assumption that every positive pool contains only one infected tick. The MIR was calculated using the formula: MIR = [(number of positive pools)/(total number of tested ticks)] × 1000 [55, 56].
Availability of data and materials
All data generated or analysed during this study are included in this published article. All the nucleotide sequences generated from this study have been deposited and are available in the GenBank database (NCBI accession No.: MW916823 and MW916824).
Abbreviations
- UR-qPCR:
-
Ultra-rapid real-time PCR
- ITS:
-
Internal transcribed spacer
- MIR:
-
Minimum infection rate
References
Fournier PE, Raoult D. Current knowledge on phylogeny and taxonomy of Rickettsia spp. Ann N Y Acad Sci. 2009;1166:1–11.
Merhej V, Angelakis E, Socolovschi C, Raoult D. Genoty**, evolution and epidemiological findings of Rickettsia species. Infect Genet Evol. 2014;25:122–37.
Parola P, Paddock CD, Socolovschi C, Labruna MB, Mediannikov O, Kernif T, et al. Update on tick-borne rickettsioses around the world: a geographic approach. Clin Microbiol Rev. 2013;26:657–702.
Parola P, Paddock CD, Raoult D. Tick-borne rickettsioses around the world: emerging diseases challenging old concepts. Clin Microbiol Rev. 2005b;18:719–56.
Borawski K, Dunaj J, Pancewicz S, Król M, Czupryna P, Moniuszko-Malinowska A. Tick-borne rickettsioses in Europe – a review. Przegl Epidemiol. 2019;73:523–30.
Oteo JA, Portillo A. Tick-borne rickettsioses in Europe. Ticks Tick Borne Dis. 2012;3:271–8.
Gaowa ON, Aochi M, Wuritu D, Wu YY, et al. Rickettsiae in ticks, Japan, 2007–2011. Emerg Infect Dis. 2013;19:338–40.
Liu H, Li Q, Zhang X, Li Z, Wang Z, Song M, et al. Characterization of rickettsiae in ticks in northeastern China. Parasit Vectors. 2016;9:498.
Shao JW, Zhang XL, Li WJ, Huang HL, Yan J. Distribution and molecular characterization of rickettsiae in ticks in Harbin area of northeastern China. PLoS Negl Trop Dis. 2020;14:e0008342.
Luce-Fedrow A, Mullins K, Kostik AP, St John HK, Jiang J, Richards AL. Strategies for detecting rickettsiae and diagnosing rickettsial diseases. Future Microbiol. 2015;10:537–64.
Merino O, De la Cruz NI, Martinez J, Pérez de León AA, Romero-Salas D, Esteve-Gassent MD, et al. Molecular detection of Rickettsia species in ticks collected in the Mexico–USA transboundary region. Exp Appl Acarol. 2020;80:559–67.
Ortega-Morales AIO, Reyna EN, Rodríguez VÁ, Álvarez VHG, Martínez AC, Rodríguez QKS, et al. Detection of Rickettsia spp. in Rhipicephalus sanguineus (sensu lato) collected from free-roaming dogs in Coahuila state, northern Mexico. Parasit Vectors. 2019;12:130.
Schattner A, Leitner M, Keysary A, Geltner D. Case report: fatal seronegative rickettsial infection diagnosed by the polymerase chain reaction. Am J Med Sci. 1992;303:392–4.
Webb L, Carl M, Malloy DC, Dasch GA, Azad AF. Detection of murine typhus infection in fleas by using the polymerase chain reaction. J Clin Microbiol. 1990;28:530–4.
Roux V, Rydkina E, Eremeeva M, Raoult D. Citrate synthase gene comparison, a new tool for phylogenetic analysis, and its application for the rickettsiae. Int J Syst Bacteriol. 1997;47:252–61.
Fournier PE, Roux V, Raoult D. Phylogenetic analysis of spotted fever group rickettsiae by study of the outer surface protein rOmpA. Int J Syst Bacteriol. 1998;48:839–49.
Roux V, Raoult D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int J Syst Evol Microbiol. 2000;50(Pt 4):1449–55.
Fournier PE, Dumler JS, Greub G, Zhang J, Wu Y, Raoult D. Gene sequence-based criteria for identification of new Rickettsia isolates and description of Rickettsia heilongjiangensis sp. nov. J Clin Microbiol. 2003;41:5456–65.
Sekeyova Z, Roux V, Raoult D. Phylogeny of Rickettsia spp. inferred by comparing sequences of ‘gene D’, which encodes an intracytoplasmic protein. Int J Syst Evol Microbiol. 2001;51(Pt 4):1353–60.
Ishikura M, Fujita H, Ando S, Matsuura K, Watanabe M. Phylogenetic analysis of spotted fever group rickettsiae isolated from ticks in Japan. Microbiol Immunol. 2002;46:241–7.
Egizi A, Gable S, Jordan RA, Rickettsia spp. Infecting lone star ticks (Amblyomma americanum) (Acari: Ixodidae) in Monmouth County, New Jersey. J Med Entomol. 2020;57:974–8.
Kato CY, Chung IH, Robinson LK, Austin AL, Dasch GA, Massung RF. Assessment of real-time PCR assay for detection of Rickettsia spp. and Rickettsia rickettsii in banked clinical samples. J Clin Microbiol. 2013;51:314–7.
Tomassone L, De Meneghi D, Adakal H, Rodighiero P, Pressi G, Grego E. Detection of Rickettsia aeschlimannii and Rickettsia africae in ixodid ticks from Burkina Faso and Somali region of Ethiopia by new real-time PCR assays. Ticks Tick Borne Dis. 2016;7:1082–8.
Kim B, Kim J, Kim S, Kim M, Truong AT, Cho K, et al. Detection of chronic bee paralysis virus using ultra-rapid PCR and nested ultra-rapid PCR. J Apic Res. 2019;58:133–40.
Kim JM, Lim SJ, Kim S, Kim M, Kim B, Truong AT, et al. Rapid detection of deformed wing virus in honeybee using ultra-rapid qPCR and a DNA-chip. J Vet Sci. 2020;21:e4.
Truong AT, Kim B, Kim S, Kim M, Kim J, Kim S, et al. Rapid detection of Israeli acute paralysis virus using multi-point ultra-rapid real-time PCR (UR-qPCR). J Apic Res. 2019;58:746–53.
Chung MH, Lee SH, Kim MJ, Lee JH, Kim ES, Lee JS, et al. Japanese spotted fever, South Korea. Emerg Infect Dis. 2006;12:1122–4.
Kim YS, Choi YJ, Lee KM, Ahn KJ, Kim HC, Klein TA, et al. First isolation of Rickettsia monacensis from a patient in South Korea. Microbiol Immunol. 2017;61:258–63.
Jang WJ, Lee MK, Kim HC, Choi YJ, Kim J, Park HJ, et al. Detection of Candidatus Rickettsia tarasevichiae from tick collected from human patient, South Korea. Syst Appl Acarol. 2019;24:193–7.
Jiang J, Choi YJ, Kim J, Kim HC, Klein TA, Chong ST, et al. Distribution of Rickettsia spp. in ticks from northwestern and southwestern provinces, Republic of Korea. Korean J Parasitol. 2019;57:161–6.
Lee JH, Ahn SJ, Park HS, Jeong EJ, Choi HG, Jang WJ, et al. Prevalence of spotted fever group Rickettsia from Haemaphysalis ticks in Chungju Province. J Bacteriol Virol. 2005;35:203–7.
Lee KM, Choi YJ, Shin SH, Choi MK, Song HJ, Kim HC, et al. Spotted fever group Rickettsia closely related to Rickettsia monacensis isolated from ticks in south Jeolla Province, Korea. Microbiol Immunol. 2013;57:487–95.
Noh Y, Lee YS, Kim HC, Chong ST, Klein TA, Jiang J, et al. Molecular detection of Rickettsia species in ticks collected from the southwestern provinces of the Republic of Korea. Parasit Vectors. 2017;10:20.
Tariq M, Seo JW, Kim DY, Panchali MJL, Yun NR, Lee YM, et al. First report of the molecular detection of human pathogen Rickettsia raoultii in ticks from the Republic of Korea. Parasit Vectors. 2021;14:191.
Jiang J, An H, Lee JS, O’Guinnd ML, Kim HC, Chong ST, et al. Molecular characterization of Haemaphysalis longicornis-borne rickettsiae, Republic of Korea and China. Ticks Tick Borne Dis. 2018;9:1606–13.
Park HJ, Kim J, Choi YJ, Kim HC, Klein TA, Chong ST, et al. Tick-borne rickettsiae in midwestern region of Republic of Korea. Acta Trop. 2021;215:105794.
Jiao J, Zhang J, He P, OuYang X, Yu Y, Wen B, et al. Identification of tick-borne pathogens and genoty** of Coxiella burnetii in Rhipicephalus microplus in Yunnan Province, China. Front Microbiol. 2021;12:736484.
Kim JY, Yi MH, Mahdi AAS, Yong TS. iSeq 100 for metagenomic pathogen screening in ticks. Parasit Vectors. 2021;14:346.
Qin XR, Han HJ, Han FJ, Zhao FM, Zhang ZT, Xue ZF, et al. Rickettsia japonica and novel Rickettsia species in ticks, China. Emerg Infect Dis. 2019;25:992–5.
Yuan TT, Du CH, **a LY, Que TC, von Fricken ME, Jiang BG, et al. Molecular evidence of Candidatus Rickettsia longicornii and a novel Rickettsia strain from ticks in southern China. Ticks Tick Borne Dis. 2021;12:101679.
Chae JB, Cho YS, Cho YK, Kang JG, Shin NS, Chae JS. Epidemiological investigation of tick species from near domestic animal farms and cattle, goat, and wild boar in Korea. Korean J Parasitol. 2019;57:319–24.
Chong ST, Kim HC, Lee IY, Kollars TM, Sancho AR, Sames WJ, et al. Seasonal distribution of ticks in four habitats near the demilitarized zone, Gyeonggi-do (province), Republic of Korea. Korean J Parasitol. 2013;51:319–25.
Zhao L, Li J, Cui X, Jia N, Wei J, **a L, et al. Distribution of Haemaphysalis longicornis and associated pathogens: analysis of pooled data from a China field survey and global published data. Lancet Planet Health. 2020;4:e320–e9.
Kim JY, Jung M, Kho JW, Song H, Moon K, Kim YH, et al. Characterization of overwintering sites of Haemaphysalis longicornis (Acari: Ixodidae) and tick infection rate with severe fever with thrombocytopenia syndrome virus from eight provinces in South Korea. Ticks Tick Borne Dis. 2020;11:101490.
Sun J, Lin J, Gong Z, Chang Y, Ye X, Gu S, et al. Detection of spotted fever group Rickettsiae in ticks from Zhejiang Province, China. Exp Appl Acarol. 2015;65:403–11.
Noden BH, Martin J, Carrillo Y, Talley JL, Ochoa-Corona FM. Development of a loop-mediated isothermal amplification (LAMP) assay for rapid screening of ticks and fleas for spotted fever group Rickettsia. PLoS One. 2018;13:e0192331.
Noden BH, Loss SR, Maichak C, Williams F. Risk of encountering ticks and tick-borne pathogens in a rapidly growing metropolitan area in the U.S. Great Plains. Ticks Tick Borne Dis. 2017;8:119–24.
Yamaguti N, Tipton VJ, Keegan HL, Toshioka S. Ticks of Japan, Korea, and the Ryukyu Islands. Brigham Young Univ Sci Bull Biol. 1971;15:1–9.
Vitorino L, Zé-Zé L, Sousa A, Bacellar F, Tenreiro R. rRNA intergenic spacer regions for phylogenetic analysis of Rickettsia species. Ann N Y Acad Sci. 2003;990:726–33.
Ishikura M, Ando S, Shinagawa Y, Matsuura K, Hasegawa S, Nakayama T, et al. Phylogenetic analysis of spotted fever group rickettsiae based on gltA, 17-kDa, and rOmpA genes amplified by nested PCR from ticks in Japan. Microbiol Immunol. 2003;47:823–32.
Regnery RL, Spruill CL, Plikaytis BD. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J Bacteriol. 1991;173:1576–89.
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23:2947–8.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98NT. Nucl Acids Symp Ser. 1999;41:95–8.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4.
Gu W, Lampman R, Novak RJ. Problems in estimating mosquito infection rates using minimum infection rate. J Med Entomol. 2003;40:595–6.
Ott D, Ulrich K, Ginsbach P, Öhme R, Bock-Hensley O, Falk U, et al. Tick-borne encephalitis virus (TBEV) prevalence in field-collected ticks (Ixodes ricinus) and phylogenetic, structural and virulence analysis in a TBE high-risk endemic area in southwestern Germany. Parasit Vectors. 2020;13:303.
Acknowledgements
The authors are thankful to Prof. Byoungsu Yoon (Kyonggi University, South Korea) for kindly supporting us.
Funding
This work was supported by the Animal and Plant Quarantine Agency [Grant number B-1543081-2020-22-03].
Author information
Authors and Affiliations
Contributions
ATT, BRY, and YSC performed study design and conceptualization. ATT, MSY, SSY, YMY, JTK, and YSC performed writing review and preparation in original draft of manuscript. ATT, JL, and SM performed experiments. ATT performed figure and table preparation. ATT and YSC performed final editing and revision. YSC performed as a laboratory supervisor, funding acquisition, and as corresponding author. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was carried out in compliance with the ARRIVE guidelines. This research project was approved by the Ethics Committee of the Animal and Plant Quarantine Agency (APQA) (Approval Number 2020–537) in the Republic of Korea. All the methods were carried out in accordance with the guidelines and regulations of Institutional Animal Care and Use Committee (IACUC) of APQA. Tick collection from wild animals was approved by Gangwon and Jeju provinces in the Republic of Korea. Ticks were collected from wild animals at the Wildlife Rescue Center with the cooperation of veterinarians, and from livestock with oral consent from all owners of animals.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Truong, AT., Yun, BR., Yoo, MS. et al. Utility of ultra-rapid real-time PCR for detection and prevalence of Rickettsia spp. in ticks. BMC Vet Res 18, 199 (2022). https://doi.org/10.1186/s12917-022-03311-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-022-03311-7