
Abstract
Background
Casuarina equisetifolia (C. equisetifolia) is a woody species with many excellent features. It has natural resistance against drought, salt and saline-alkali stresses. WRKY transcription factors (TFs) play significant roles in plant response to abiotic stresses, therefore, molecular characterization of WRKY gene family under abiotic stresses holds great significance for improvement of forest trees through molecular biological tools. At present, WRKY TFs from C. equisetifolia have not been thoroughly studied with respect to their role in salt and saline-alkali stresses response. The current study was conducted to bridge the same knowledge gap.
Results
A total of 64 WRKYs were identified in C. equisetifolia and divided into three major groups i.e. group I, II and III, consisting of 10, 42 and 12 WRKY members, respectively. The WRKY members in group II were further divided into 5 subgroups according to their homology with Arabidopsis counterparts. WRKYs belonging to the same group exhibited higher similarities in gene structure and the presence of conserved motifs. Promoter analysis data showed the presence of various response elements, especially those related to hormone signaling and abiotic stresses, such as ABRE (ABA), TGACG (MeJA), W-box ((C/T) TGAC (T/C)) and TC-rich motif. Tissue specific expression data showed that CeqWRKYs were mainly expressed in root under normal growth conditions. Furthermore, most of the CeqWRKYs were up-regulated by NaCl and NaHCO3 stresses with few of WRKYs showing early responsiveness to both stresses while few others exhibiting late response. Although the expressions of CeqWRKYs were also induced by cold stress, the response was delayed compared with other stresses. Transgenic C. equisetifolia plants overexpressing CeqWRKY11 displayed lower electrolyte leakage, higher chlorophyll content, and enhanced tolerance to both stresses. The higher expression of abiotic stress related genes, especially CeqHKT1 and CeqPOD7, in overexpression lines points to the maintenance of optimum Na+/K+ ratio, and ROS scavenging as possible key molecular mechanisms underlying salt stress tolerance.
Conclusions
Our results show that CeqWRKYs might be key regulators of NaCl and NaHCO3 stresses response in C. equisetifolia. In addition, positive correlation of CeqWRKY11 expression with increased stress tolerance in C. equisetifolia encourages further research on other WRKY family members through functional genomic tools. The best candidates could be incorporated in other woody plant species for improving stress tolerance.
Similar content being viewed by others

Introduction
WRKY is one of the biggest transcription factor (TF) family in plants like Arabidopsis thaliana [1], Oryza sativa [2], Tritipyrum [3], and Isatis indigotica [4]. WRKY proteins have highly conserved structure, containing one or two WRKY domains composed of approximately 60 amino acid residues. The N-terminal of the domain contains the WRKYGQK sequence, which is related to DNA binding activity, while the C-terminal of the domain includes the C-X4-5-C-X22-23-H-X1-H or C-X7-C-X23-H-X1-C zinc finger, which participates in protein interaction and improves DNA binding [4, 5].
WRKY transcription factors play important roles in regulating plant growth, development, and response to drought, temperature and salt stresses [6, 7]. For example, OsWRKY45 plays a positive role, whereas both OsWRKY5 and OsWRKY114 separately play negative roles in response to drought stress [8,9,10]. Overexpression of BdWRKY36 and PheWRKY86 improved drought tolerance in tobacco and rice, respectively [11, 12]. Additionally, it has been demonstrated that KoWRKY40 and LlWRKY22 participate in response to temperature stress [13, 14]. At present, a large number of salt responsive WRKY transcription factors have been identified in various plants. Overexpression of FcWRKY40 in tobacco, and overexpression of VvWRKY30 and GmWRKY49 in A. thaliana confer salt tolerance in transgenic plants [15,32].
Putative promoter cis-acting element analysis and co-expression networks structure
In order to study the regulatory mechanism of the CeqWRKYs in response to abiotic stresses, the 2, 000 bp upstream region from the translation start site of the CeqWRKY genes was obtained from the C. equisetifolia database and cis-acting elements were identified by using the PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) and PlantPAN 3.0 program (PlantPAN 3.0 (ncku.edu.tw)) [42, 43]. Subsequently, we chose 11 representative cis-acting elements associated with plant growth and development, hormone response, and environmental stress responses for further analysis.
Through SPSS software, One-way analysis of variance (ANOVA) was used to analyze the Pearson correlation coefficients of the WRKYs expression data, and to further construct the co-expression networks. These networks were visualized by Cytoscape (v3.7.2) [44].
Subcellular localization and transactivation activity assay
The subcellular localization of putative CeqWRKY proteins was predicted using online website WoLP PSORT (https://wolfpsort.hgc.jp/). The coding sequences of CeqWRKYs (without the stop codon) were PCR amplified using the primers (Table S6) and then incorporated into the pCAMBIA1300-35S-GFP vector upstream of green fluorescent protein (GFP) gene. The recombinant plasmids (pCAMBIA1300-35S-CeqWRKYs::GFP) were separately transformed to the Agrobacterium tumefaciens strain GV3101. The suspension containing recombinant plasmid was injected into the lower epidermis of tobacco leaves and labeled. After two days culture in dark, GFP fluorescence was observed by confocal microscope (LSM710, Carl Zeiss, Jena, Germany).
For transactivation activity analysis, the open reading frames (ORFs) of CeqWRKYs were amplified using the primers given in Table S6 and were inserted into the pGBKT7 vector (Clontech) containing the GAL4 DNA-binding domain (BD), to get fusion vectors designated as pGBKT7-CeqWRKYs. The pGAL4 and the pGBKT7 empty vector were used as a positive and negative control, respectively. The control and fusion plasmids were separately transformed into the yeast strain AH109 according to the method of Sun et al. [45]. The transformed cells were streaked on SD/-Trp and SD/-Trp/-His media. The transactivation activity of the CeqWRKY proteins was assessed based on the growth of the yeast cells at 30℃ for 3–5 days. Three biological replicates were performed.
Generation of transgenic C. equisetifolia with overexpressing CeqWRKY11 by using efficient hairy root transgenic system
The full-length coding sequence (CDS) of CeqWRKY11 was cloned into the pCAMBIA1300-GFP vector containing the CaMV 35S promoter fused with the green fluorescent protein (GFP) reporter gene and transferred into one month old C. equisetifolia seedlings by using hairy root transgenic system [30]. The transgenic positive seedlings were identified through GFP detection by using LUYOR-3415RG (Hand-held Lamp) and the expression analysis of CeqWRKY11 by qRT-PCR. The primers used were in Table S5.
Statistical analysis
The significant difference between mean values was determined using T-test by GraphPad Prism8. Different number of asterisks against the error bars of histograms represents significant differences relative to controls, which were indicated at **P < 0.01 and *P < 0.05.
Results
Identification of WRKY genes in C. equisetifolia
The candidate genes with typical WRKY or WRKY-like domains were preliminarily screened from C. equisetifolia genomic database according to the Hidden Markov Model (HMM) profile of the WRKY domain. After removing the incomplete domain sequences, a total of 64 WRKY genes were identified in C. equisetifolia, and were named according to evolutionary relationship with those of A. thaliana genes. As shown in Table S1, the lengths of CeqWRKY protein sequences ranged from 131 to 788 amino acids, and molecular weight ranged from 14.56 KDa to 84.76 KDa. Moreover, the theoretical isoelectric point (pI) varied from 4.77 to 9.80 (Table S1). According to the predicted subcellular localization data, most of the CeqWRKY proteins were localized in the nucleus, whereas two (CeqWRKY10 and CeqWRKY70) were localized in both nucleus and cytoplasm (Table S1).
Phylogenetic trees and group classification of CeqWRKY proteins
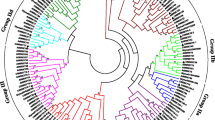
According to the alignment, the phylogenetic tree was constructed using CeqWRKYs and AtWRKYs through the MEGA7 software (Fig. 1). As shown in Fig. 1, 64 CeqWRKYs were clustered into three major groups, including group I (10 CeqWRKYs), group II (42 CeqWRKYs), and group III (12 CeqWRKYs). In Group II, CeqWRKY members were further divided into 5 subgroups, including group IIa (4 CeqWRKYs), group IIb (8 CeqWRKYs), group IIc (16 CeqWRKYs), group IId (7 CeqWRKYs) and group IIe (7 CeqWRKYs). The number of CeqWRKY distributed in different groups in C. equisetifolia was in accordance with those in A. thaliana. The largest subgroup was group IIc, while the smallest was IIa.

Phylogenetic tree showing evolutionary among WRKY domains of C. equisetifolia and A. thaliana. The phylogenetic tree was constructed using the neighbor-joining method in the MEGA7 software
Protein motifs and gene structure analysis
To further analyze the conserved domains of the CeqWRKYs, the MEME online program (http://meme-suite.org/tools/meme) was performed. Overall, 20 conserved motifs were identified in CeqWRKYs; the sequences of these motifs were shown in Table S2. Moreover, we found that CeqWRKY from the same group or subgroup contained similar conserved motifs. In detail, Motif 1 and Motif 3 consisted of the conserved heptapeptide domains (WRKYGQK) of WRKY proteins (Fig. 2A, Table S2). Motif 1 and Motif 2 were distributed in most of the WRKYs, while Motif 3 was only distributed in group I. Motif 8 was unique in group IIa and IIb (Fig. 2A). Although the functions of the conserved motifs have not yet been understood, the classification based on the conserved motifs of the CeqWRKY proteins has widely been used.

Phylogenetic relationships, conserved motifs and gene structure analysis of CeqWRKYs. A Phylogenetic tree and conserved motifs of CeqWRKYs. Different colored boxes represent the presence of 1–20 conserved motifs. B Gene structure analysis of CeqWRKYs. Introns are depicted by black lines, exons by green boxes, and untranslated 5’- and 3’-regions by yellow boxes
As the evolution of multi-gene families is attributed to the diversity of the gene structure, so the exon–intron structure of the 64 CeqWRKY genes was analyzed. The exons in genes from group I ranged from 4–7 while those in group IIa, group IIb, and group IIc ranged from 4–5, 4–8, and 2–5, respectively (Fig. 2B). All the members of group IId and most of group IIe and group III contained three exons (Fig. 2B). Genes in the same group generally had similar structures, but significant differences existed in the number of introns and exons in the different groups, indicating that the structure of CeqWRKY genes was relatively complex.
The analysis of cis-acting elements in CeqWRKY gene promoters
In order to study the potential regulatory role of CeqWRKY transcription factors in stress responses, the cis-acting regulatory elements contained in the promoters of 64 CeqWRKY genes were analyzed. The results showed that all CeqWRKY genes contained diverse cis-acting elements in their promoter regions. Further, the distribution of 11 representative cis-acting regulatory elements in promoter regions was investigated. Based on their functions, these cis-acting elements were divided into three categories: category 1 included motifs related to hormone response (ABRE, P-box, TATC-box, TGACG-motif, AuxRR-core, TGA-element), and category 2 had motifs related to environmental stress (WUN-motif, TC-rich repeats, LTR and MBS), while category 3 included motifs related to development (MBSI) [32, 46, 47]. The analysis showed that abundant cis-acting elements were distributed in the promoter regions of CeqWRKYs (Fig. 3A). More than half of CeqWRKYs contained 3–5 ABRE elements (Fig. 3B), which play important roles in ABA-mediated diverse stress response in plants [47]. In addition, 52 CeqWRKYs contained TGACG-motifs that could be related to MeJA response pathway. Additionally, 34 CeqWRKYs containing P-box and 11 CeqWRKYs containing TATC-box may be involved in gibberellin response, while 8 CeqWRKYs containing AuxRR-core and 3 CeqWRKYs containing TGA-element were related to auxin response (Fig. 3B). The promoters of CeqWRKY genes also contained a substantial number of environmental stress-related response elements, with 24 CeqWRKYs containing TC-rich repeats (Fig. 3B) and all CeqWRKYs containing W-box (Fig. S1). Only CeqWRKY35 and CeqWRKY49 included WUN-motif (wound-responsive element), which may play critical roles in plant response to mechanical damage. Additionally, many CeqWRKYs’ promoters included LTR (low-temperature stress responsive element) and MBS (drought stress responsive element) which significantly contribute to plant responses to low temperature and drought stress.

Cis-elements analysis in the CeqWRKYs promoter region. A The number of different types of cis-elements in the CeqWRKYs promoter region. B The number of 11 representative cis-acting regulatory elements in promoter regions of CeqWRKYs
Determination of expression pattern analysis of CeqWRKYs under cold, NaCl, NaHCO3 and pH stresses
To further investigate the changes in CeqWRKYs gene expression under different abiotic stresses, the transcriptome data of C. equisetifolia under four different treatments i.e. 300 mM NaCl, 300 mM NaHCO3, pH 8.5 and low temperature (4℃), at different time points were analyzed (Fig. 4, Tables S3 and S4). 43 CeqWRKYs were highly expressed in root, while 13 CeqWRKYs showed strong expression in shoot (Fig. 4A). Under cold treatment, the majority of CeqWRKYs displayed increased expression as treatment time increased (Fig. 4B). CeqWRKY10 and CeqWRKY44 were strongly up-regulated at 10 min and 2 h time points under low temperature stress. 28 CeqWRKYs were significantly induced by cold treatment at 168 h time point (Fig. 4B). According to the expression patterns of WRKY genes under NaCl, NaHCO3, and pH stresses, we found that some CeqWRKYs responded to two stresses. For example, in shoot, CeqWRKY6, CeqWRKY17, CeqWRKY22, CeqWRKY31, CeqWRKY46, CeqWRKY50, and CeqWRKY60 were induced by both NaCl and NaHCO3 stresses, while CeqWRKY36, CeqWRKY40, CeqWRKY48, CeqWRKY51, and CeqWRKY72 were up-regulated by both NaHCO3 and pH stresses (Fig. 4C). In root, plenty of CeqWRKYs also displayed much higher expression than control under different stresses, such as CeqWRKY11, CeqWRKY33, and CeqWRKY41 under NaCl stress, CeqWRKY39, CeqWRKY46, and CeqWRKY71 under NaHCO3 stress, and CeqWRKY14, CeqWRKY50, and CeqWRKY70 under pH stress (Fig. 4D). These results indicated that WRKYs could be involved in diverse abiotic stresses response in C. equisetifolia.

Heatmap of CeqWRKYs expression using RNA-seq data. Blue color represents low expression levels, while red color represents high expression levels. A The expression patterns of CeqWRKYs in root and shoot. B The expression of CeqWRKYs in shoot under cold stress. C-D The expression patterns of CeqWRKYs in shoot and root under NaCl, NaHCO3, and pH stresses
Identification of CeqWRKYs expression in root and shoot under NaCl and NaHCO3 stresses
Previous study has reported that C. equisetifolia has strong salt and saline-alkali tolerance [48]. To verify the transcriptional changes and identify the candidate CeqWRKYs involved in response to NaCl and NaHCO3 conditions, RNA was extracted from the shoot and root of C. equisetifolia grown under control, NaCl and NaHCO3 treatment at different time points, respectively, and the quality of total RNA was detected (Table S7). Then, the expression of 60 CeqWRKYs in root and shoot was determined by qRT-PCR at different time points.
Under salt treatment, some CeqWRKYs such as WRKY18, WRKY29, WRKY33, WRKY40, WRKY41, displayed significant up-regulation at 0.5 h after the start of the salt treatment, while some CeqWRKYs such as WRKY35, WRKY42, WRKY50, and WRKY64 were induced at 24 h in root (Fig. 5A). CeqWRKY4, CeqWRKY10, CeqWRKY33, CeqWRKY38, CeqWRKY40, CeqWRKY53, and CeqWRKY62 displayed rapid and sustained response to salt stress in root (Fig. 5A). In shoot, some salt-responsive CeqWRKYs, including CeqWRKY11, CeqWRKY17, CeqWRKY22, CeqWRKY25, CeqWRKY33, CeqWRKY40, CeqWRKY41, CeqWRKY46, CeqWRKY53, CeqWRKY62, CeqWRKY70, and CeqWRKY77, showed different degrees of induced expression (Fig. 5B). In addition, CeqWRKY25 and CeqWRKY40 showed a persistent high expression at different time points under salt stress in both root and shoot (Fig. 5).

Expression profiles of CeqWRKYs under NaCl stress. A The expression patterns of CeqWRKYs under NaCl stress in root. B The expression patterns of CeqWRKYs under NaCl stress in shoot. The mean ± standard error measurement (SEM) value with three replications is displayed. Relative expression in untreated plants (0 h) was set to 1. T-tests were used to indicate significant difference: * indicates p < 0.05, and ** indicates p < 0.01
Under NaHCO3 stress, many CeqWRKYs showed significant early response (0.5 h) and late response (12 h or 24 h). In roots, CeqWRKY11, CeqWRKY18, CeqWRKY33, CeqWRKY34, CeqWRKY41, CeqWRKY46, CeqWRKY64, and CeqWRKY70 were obviously up-regulated at early treatment time point, while CeqWRKY15, CeqWRKY28, CeqWRKY48, CeqWRKY65 and CeqWRKY77 responded to the NaHCO3 stress at late time points (Fig. 6A). In shoot, there were some early responsive CeqWRKY genes, such as CeqWRKY17, CeqWRKY18, CeqWRKY22, CeqWRKY25, CeqWRKY33, CeqWRKY40, CeqWRKY41, CeqWRKY46, CeqWRKY53, and CeqWRKY61 (Fig. 6B). Compared to the expression pattern of CeqWRKYs in root, no CeqWRKY genes displayed significant late response to NaHCO3 stress in shoot (Fig. 6B). Additionally, some CeqWRKYs maintained continuous induction under NaHCO3 stress, such as CeqWRKY4, CeqWRKY25, and CeqWRKY62 in root, and CeqWRKY17, CeqWRKY22, CeqWRKY28, CeqWRKY29, CeqWRKY31, CeqWRKY39, CeqWRKY61, and CeqWRKY65 in shoot (Fig. 6).

Expression profiles of CeqWRKYs under NaHCO3 stress. A The expression patterns of CeqWRKYs under NaHCO3 stress in root. B The expression patterns of CeqWRKYs under NaHCO3 stress in shoot. The mean ± standard error measurement (SEM) value with three replications is displayed. Relative expression in untreated plants (0 h) was set to 1. T-tests were used to indicate significant difference: * indicates p < 0.05, and ** indicates p < 0.01
Since salinization and alkalization often occur simultaneously in natural soil environments [49], it was important to identify a set of genes induced by both NaCl and NaHCO3 stresses. To this end, we identified 20 CeqWRKYs responsive to both stresses, such as CeqWRKY11, CeqWRKY28, CeqWRKY29, CeqWRKY31, CeqWRKY33, CeqWRKY41, CeqWRKY45, CeqWRKY46, and CeqWRKY50, etc. (Figs. 5 and 6). In addition, some CeqWRKY genes displayed different response under NaCl and NaHCO3 stresses. For instance, CeqWRKY21, CeqWRKY23, and CeqWRKY47 were only involved in response to NaHCO3 stress both in root and shoot, but not to NaCl stress (Figs. 5 and 6).
Co-expression networks analysis of CeqWRKYs
WRKY TFs regulate the expression of downstream target genes by binding to W-box located in their promoter region [50]. Due to the presence of different number of W-box elements in the CeqWRKYs’ promoter (Fig. S1). Co-expression networks analysis was performed for all CeqWRKYs using transcription expression data under NaCl and NaHCO3 stresses in both root and shoot. The co-expression networks data showed that there were complex regulation networks among CeqWRKYs and that one CeqWRKY could regulate several other CeqWRKYs (Fig. 7).

Co-expression networks among CeqWRKYs based on the gene expression in transcriptome data. A-B Co-expression networks among CeqWRKYs under NaCl stress in root and shoot, respectively. C-D Co-expression networks among CeqWRKYs under NaHCO3 condition in root and shoot, respectively. Degree value represents the number of connecting lines around a node (circle). Larger and darker coloured circles indicate strong correlation with other genes. Edges are drawn when the linear correlation coefficient is > 0.8 with p-value < 0.05
Under NaCl stress, there were several WRKYs with higher degree values at the core in the larger network of root, such as CeqWRKY41, CeqWRKY11, CeqWRKY29, CeqWRKY62, CeqWRKY40 and CeqWRKY33. In the smaller network, CeqWRKY34 may take key parts in response to salt stress in root (Fig. 7A). In shoot, the putative co-expression network showed relatively dispersed, compared to that in root, and the CeqWRKY genes located in the center position of each network was different, including CeqWRKY41, CeqWRKY31, and CeqWRKY35 (Fig. 7B). These indicated that CeqWRKYs could form different regulatory networks through interaction with other CeqWRKYs under salt stress response.
Under NaHCO3 stress, CeqWRKY41, CeqWRKY46, CeqWRKY60, CeqWRKY11, CeqWRKY31, CeqWRKY48, and CeqWRKY33 may function as the key regulators to mediate NaHCO3 stress response in root (Fig. 7C). In shoot, the core genes were same as those in root, but had higher value in shoot than in root. Besides that, CeqWRKY1 and CeqWRKY35 also had higher degree value in shoot (Fig. 7D).
The analysis of subcellular localization and transcriptional activation activity of CeqWRKYs
Previous studies showed that WRKY11, WRKY33, WRKY41, and WRKY46 from different plant species play key roles in response to diverse stresses [47,48,49]. According to the co-expression networks, these four CeqWRKYs may play important roles in response to NaCl and NaHCO3 stress. To verify whether these WRKYs function as transcription factors in C. equisetifolia, the subcellular localization of the above mentioned WRKYs was analyzed. The constructs (35S-CeqWRKYs::GFP) of these genes were separately transformed into tobacco leaves by Agrobacterium tumefaciens-mediated transient method. Compared to the empty vector control, the GFP signal from different CeqWRKYs fused proteins was detected only in the nucleus (Fig. 8A), indicating that CeqWRKY11, CeqWRKY33, CeqWRKY41 and CeqWRKY46 were specifically nucleus-localized proteins, and may function as TFs in stress responses.

Subcellular localization and transcriptional activation activity analysis. A Subcellular localization analysis of CeqWRKY11, CeqWRKY33, CeqWRKY41, and CeqWRKY46. B Transcriptional activation activity analysis of CeqWRKY11, CeqWRKY33, CeqWRKY41, and CeqWRKY46
To further analyze the transcriptional activation activity of these WRKYs, the pGBKT7-CeqWRKYs plasmids (for CeqWRKY11, CeqWRKY33, CeqWRKY41 and CeqWRKY46) were transformed into AH109 yeast cells. pGAL4 and empty vector pGBKT7 were used as positive and negative control, respectively. All the transformants exhibited growth on the SD/-Trp plate (Fig. 8B). Moreover, the transformants carrying the pGBKT7-CeqWRKYs plasmids and the positive control grew well on SD/-Trp/-His medium, whereas the negative control did not grow (Fig. 8B). These data demonstrated that the CeqWRKYs could exhibit remarkable self-activation activity.
Phenotype analysis of transgenic C. equisetifolia plants with overexpressing CeqWRKY11 under NaCl and NaHCO3 stresses
To further study the function of CeqWRKY11 in NaCl and NaHCO3 response, transgenic C. equisetifolia plants with overexpressing CeqWRKY11 were developed by hairy-root mediated transgenic method, and their phenotype was analyzed under 200 mM NaCl and 75 mM NaHCO3 conditions, respectively. Considering the fusion of CeqWRKY11 and GFP protein in recombinant plasmid, the GFP fluorescence was detected to identify the transgenic positive C. equisetifolia plants. The results showed that, compared to the negative control (WT), there was obvious green fluorescence in overexpressing CeqWRKY11 lines and the positive control (with empty overexpressing vector) (Fig. S3). The expression of CeqWRKY11 in transgenic line roots (WRKY11OE-1, WRKY11OE-2) was confirmed by qRT-PCR, and was found to be higher than control plants (CK: transformed with empty vector) (Fig. 9E), demonstrating the overexpression of CeqWRKY11 in transgenic positive lines. Under normal growth condition, no significant difference was observed in the growth of transgenic plants compared to control (Fig. 9A, C). However, after 5 days of NaCl treatment, the shoots of the CK were severely wilted, but the shoots of overexpression lines grew normally and displayed much greener (Fig. 9B). When grown under NaHCO3 treatment for 15 days, the roots of the CK plants showed severe browning, but WRKY11OE displayed normal growth without significant changes (Fig. 9D). In addition, there was more abundant chlorophyll content in shoots of the overexpression lines than in control plants under NaCl condition, while there was no significant difference in chlorophyll content between control plants and WRKY11 overexpression plants under NaHCO3 treatment (Fig. 9F). After NaCl or NaHCO3 treatment, although the electrolyte leakage was obviously increased in both the overexpression lines and control plants, but compared to control plants, the overexpression lines showed much lower electronic leakage (Fig. 9G). Taken together, these results indicated that the expression of CeqWRKY11 improves the tolerance of C. equisetifolia to NaCl and NaHCO3 stresses.

Phenotype analysis of CeqWRKY11-overexpressing transgenic plants (CeqWRKY11OE-1, CeqWRKY11OE-2) in C.equisetifolia under NaCl and NaHCO3 condition. A-B Phenotype analysis of CeqWRKY11-overexpressing transgenic plants under normal and 200 mM NaCl condition. C-D Phenotype analysis of CeqWRKY11-overexpressing transgenic plants under normal and 75 mM NaHCO3 condition. E The expression of CeqWRKY11 in control and CeqWRKY11-overexpressing transgenic plants. F Chlorophyll content in control and CeqWRKY11-overexpressing transgenic plants under normal, 200 mM NaCl and 75 mM NaHCO3 condition. G Electronic leakage of control and CeqWRKY11-overexpressing transgenic plants under normal, 200 mM NaCl and 75 mM NaHCO3 condition. T-tests were used to indicate significant difference: * indicates p < 0.05, and ** indicates p < 0.01
Overexpression of CeqWRKY11 altered the expression of stress-responsive genes in C. equisetifolia
To investigate the molecular mechanism of higher NaCl and NaHCO3 stress tolerance of CeqWRKY11 overexpressing lines, we analyzed the expression of several stress-responsive genes (CeqHKT1, CeqNHX, CeqRD29A, CeqUGTs), and oxidative stress related genes (CeqPOD, CeqRbohD, CeqCSD1, CeqGols) in the CeqWRKY11 overexpression lines and control plants under normal condition. The results showed that all these genes, especially CeqHKT1 and CeqPOD7, were markedly up-regulated in transgenic lines compared with control (Fig. 10), indicating that the overexpression of CeqWRKY11 may improve the NaCl and NaHCO3 tolerance in C. equisetifolia through regulating the Na+/K+ balance and maintaining a stable ROS level.

Expression analysis of stress-responsive genes in CeqWRKY11 overexpression (OE-WRKY11) and non-transgenic C. Equisetifolia (CK) plants by qRT-PCR. The mean ± standard error measurement (SEM) value with three replications is displayed. Relative expression in CK was set to 1. T-tests were used to indicate significant difference: * indicates p < 0.05, and ** indicates p < 0.01
Discussion
The WRKY gene family, one of the largest transcription factor families, plays essential roles in various aspects of plant development and their ability to respond to both biotic and abiotic stresses [5, 51]. Previous studies have been identified WRKY genes in different plant species, including 75 WRKYs in A. thaliana [52], 126 in O. sativa [53], 81 in Solanum lycopersicum [54], 48 in Camellia japonica [55], 197 in Glycine max [56], and 98 in Populus trichocarpa [57]. In this study, a total of 64 WRKY genes were identified in C. equisetifolia (Fig. 1), which was smaller, compared to the number of WRKY members in other species. The CeqWRKY members were categorized into three major groups; group II was further divided into five subgroups (Fig. 2). Furthermore, the distribution of CeqWRKY genes within each group or subgroup was similar to that of A. thaliana and P. trichocarpa [1, 57]. Previous study has demonstrated that genes expression and function were highly related to their regulatory elements in promoter [47]. Our study found that the promoter region of group IIc CeqWRKYs included more stress-related cis-elements (Fig. 3), and the expression of most of the CeqWRKYs was induced by at least one stress treatment (Figs. 5 and 6), indicating that group IIc CeqWRKYs could be involved in mediating abiotic stress resistance in C. equisetifolia.
Plants respond to many environmental stresses by regulating the complex signal transduction pathways. The cis-element analysis showed that, compared to the promoter region of WRKY genes from Taraxacum kok-saghyz and Liriodendron chinense, CeqWRKYs promoter region contained much more hormone response elements, including ABRE, MeJA, and stress related elements, such as W-box (WRKY transcription factor binding site), and TC-rich (salt stress-related elements) (Fig. 3B, Figure S1) [46, 58], inferring that CeqWRKYs are involved in ABA- and MeJA-mediated stress responses.
Gene expression is highly correlated with gene function [59]. Previous studies showed that genes specifically expressed in the roots were assumed to be key regulators of root development and may play roles in response to various stresses [60, 61]. Our results showed that many CeqWRKYs were highly expressed under normal condition in root or shoot, implying a constitutive expression for WRKYs in C. equisetifolia (Fig. 4A), which may be in accordance with the strong inherent stress resistance of C. equisetifolia. In addition, we found that CeqWRKYs quickly and continuously responded to NaCl and NaHCO3 stress, but delayed responded to cold stress (Fig. 4B, C, D), which may be one of the reasons for its sensitivity to cold stress. The latest research showed that WRKYs are mainly involved in response to salt stress [16]. In our study, we found that most of CeqWRKYs genes were separately responsive to NaCl or NaHCO3 stress at the early and late treatment time points, and some CeqWRKYs displayed continuous response (Figs. 5 and 6). Furthermore, there were different CeqWRKYs independently involved in NaCl or NaHCO3 stress. For instance, CeqWRKY47 was only induced by NaHCO3 stress, while CeqWRKY24 was only up-regulated by NaCl stress (Figs. 5 and 6). In addition, the induced expression of CeqWRKYs also displayed tissue specificity. Under NaCl stress, CeqWRKY17 was only up-regulated in shoot while CeqWRKY10 and CeqWRKY34 were only induced in root (Fig. 5). These results illustrated that CeqWRKYs displayed tissue specificity in expression pattern and functional diversification in response to salt stress. Notably, some CeqWRKYs respond to NaCl and NaHCO3 stresses at the same time. For example, CeqWRKY11, 33, 41, 46 were significantly up-regulated by NaCl and NaHCO3 stresses in both root and shoot. Moreover, these four CeqWRKYs were identified as core genes in co-expression networks, implying that these CeqWRKYs may play more important role in NaCl and NaHCO3 stress response (Figs. 5, 6 and 7). Furthermore, all four CeqWRKYs were localized in nucleus and showed self-transcriptional activation activity (Fig. 8). In addition, comparatively less electrolyte leakage and higher chlorophyll content in the CeqWRKY11 overexpression lines than control under NaCl and NaHCO3 treatment condition, respectively (Fig. 9), showed that CeqWRKY11 may play a positive role in NaCl and NaHCO3 stress response in C. equisetifolia, which are similar to the functional characterization of Arabidopsis WRKY11 [62]. Previous study has reported that, under salt stress, C. equisetifolia can impede Na+ transfer to the shoot through sequestering Na+ in root tissue via Na+ and K+ transport proteins, and the expression of genes related to oxidative stress and detoxification was up-regulated [63, 64]. In our study, the expression of CeqHKT1 and CeqPOD7 in overexpressing WRKY11 transgenic plants was marked increased (Fig. 10), indicating that C. equisetifolia response to NaCl and NaHCO3 stresses could partly involve in HKT1-mediated Na+ and K+ transport and POD-mediated ROS scavenging activity.
Transcriptional regulation plays significant role in plants response to multiple stresses [65]. Under diverse stresses, different TFs were activated and then immediately induced the expression of the downstream stress-responsive genes, which triggers a set of responses to minimize stress damage. Recent study showed that OsWRKY63-OsWRKY76-OsDREB1B transcriptional regulatory cascade plays key role in chilling tolerance of rice [66]. Based on the CeqWRKYs’ co-expression and interaction networks data (Fig. 7, Fig. S2), we hypothesize that CeqWRKY may interact with each other CeqWRKY(s) or form a transcription cascades to regulate the expression of downstream genes to respond to diverse stresses.
Conclusions
We identified 64 WRKY genes in C. equisetifolia and divided into three major groups. Group II was further divided into 5 subgroups. The CeqWRKY genes were highly similar with the conserved domains. Their promoter regions contained various response elements. According to the gene expression analysis, WRKYs showed delayed transcriptional response to cold stress while a rapid response with varying degrees to NaCl and NaHCO3 stresses. CeqWRKYs may take important parts in response to NaCl and NaHCO3 stresses through the interaction among CeqWRKYs. Furthermore, overexpression of CeqWRKY11 enhanced the tolerance of transgenic C. equisetifolia seedlings to NaCl and NaHCO3 stress by up-regulation the expression of abiotic stress related genes, especially CeqHKT1 and CeqPOD7. This systematic analysis of CeqWRKY gene family will provide a foundation for the function and regulatory mechanism of the WRKYs in abiotic stress response and a theoretical basis for forest trees improvement.
Availability of data and materials
All data analyzed during this study are included in this article and its additional files.
References
Wang Q, Wang M, Zhang X, et al. WRKY gene family evolution in Arabidopsis thaliana. Genetica. 2011;139(8):973–83.
Ramamoorthy RR, Jiang SY, Kumar N, et al. A comprehensive transcriptional profiling of the WRKY gene family in rice under various abiotic and phytohormone treatments. Plant Cell Physiol. 2008;49(6):865–79.
Li K, Liu X, He F, et al. Genome-wide analysis of the Tritipyrum WRKY gene family and the response of TtWRKY256 in salt-tolerance. Front Plant Sci. 2022;13:1042078.
Qu R, Cao Y, Tang X, et al. Identification and expression analysis of the WRKY gene family in Isatis indigotica. Gene. 2021;783:145561.
Jiang J, Ma S, Ye N, et al. WRKY transcription factors in plant responses to stresses. J Integr Plant Biol. 2017;59(2):86–101.
Price L, Han Y, Angessa T, et al. Molecular pathways of WRKY genes in regulating plant salinity tolerance. Int J Mol Sci. 2022;23(18):10947.
Goyal P, Devi R, Verma B, et al. WRKY transcription factors, evolution, regulation, and functional diversity in plants. Protoplasma. 2023;260(2):331–48.
Qiu Y, Yu D. Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance in Arabidopsis. Environ Exp Bot. 2009;65(1):35–47.
Lim C, Kang K, Shim Y, et al. Inactivating transcription factor OsWRKY5 enhances drought tolerance through abscisic acid signaling pathways. Plant Physiol. 2022;188(4):1900–16.
Song G, Son S, Lee KS, et al. OsWRKY114 negatively regulates drought tolerance by restricting stomatal closure in rice. Plants (Basel). 2022;11(15):1938.
Sun J, Hu W, Zhou R, et al. The Brachypodium distachyon BdWRKY36 gene confers tolerance to drought stress in transgenic tobacco plants. Plant Cell Rep. 2015;34(1):23–35.
Wu M, Zhang K, Xu Y, et al. The moso bamboo WRKY transcription factor, PheWRKY86, regulates drought tolerance in transgenic plants. Plant Physiol Biochem. 2022;170:180–91.
Fei J, Wang YS, Cheng H, et al. The kandelia obovata transcription factor KoWRKY40 enhances cold tolerance in transgenic Arabidopsis. BMC Plant Biol. 2022;22(1):274.
Wu Z, Li T, Cao X, et al. Lily WRKY factor LlWRKY22 promotes thermotolerance through autoactivation and activation of LlDREB2B. Hortic Res. 2022;9:uhac186.
Dai W, Wang M, Gong X, et al. The transcription factor FcWRKY40 of Fortunella crassifolia functions positively in salt tolerance through modulation of ion homeostasis and proline biosynthesis by directly regulating SOS2 and P5CS1 homologs. New Phytol. 2018;219(3):972–89.
Zhu D, Hou L, **ao P. VvWRKY30, a grape WRKY transcription factor, plays a positive regulatory role under salinity stress. Plant Sci. 2019;280:132–42.
Xu Z, Raza Q, Xu L, et al. GmWRKY49, a salt-responsive nuclear protein, improved root length and governed better salinity tolerance in transgenic Arabidopsis. Front Plant Sci. 2018;9:809.
Fang X, Li W, Yuan H, et al. Mutation of ZmWRKY86 confers enhanced salt stress tolerance in maize. Plant Physiol Biochem. 2021;167:840–50.
Zhang M, Zhao R, Wang H, et al. OsWRKY28 positively regulates salinity tolerance by directly activating OsDREB1B expression in rice. Plant Cell Rep. 2023;42(2):223–34.
Huang J, Liu F, Chao D, et al. The WRKY transcription factor OsWRKY54 is involved in salt tolerance in rice. Int J Mol Sci. 2022;23(19):11999.
Niu Y, Li X, Xu C, et al. Analysis of drought and salt-alkali tolerance in tobacco by overexpressing WRKY39 gene from Populus trichocarpa. Plant Signal Behav. 2021;16(7):1918885.
Wang W, Bai XD, Chen K, et al. PsnWRKY70 negatively regulates NaHCO3 tolerance in populus. Int J Mol Sci. 2022;23(21):13086.
Chen F, Hu Y, Vannozzi A, et al. The WRKY transcription factor family in model plants and crops. Crit Rev Plant Sci. 2018;36(5–6):311–35.
Wu TY, Goh H, Azodi CB, et al. Evolutionarily conserved hierarchical gene regulatory networks for plant salt stress response. Nat Plants. 2021;7(6):787–99.
Bo C, Cai R, Fang X, et al. Transcription factor ZmWRKY20 interacts with ZmWRKY115 to repress expression of ZmbZIP111 for salt tolerance in maize. Plant J. 2022;111(6):1660–75.
Tani C, Sasakawa H. Salt tolerance of Casuarina equisetifolia and Frankia Ceq1 strain isolated from the root nodules of C. equisetifolia. Soil Sci Plant Nutr. 2003;49(2):215–22.
Pinyopusarerk K, Williams ER, Luangviriyasaeng V, et al. Variations in growth and morphological characteristics of Casuarina junghuhniana provenances grown in Thailand. J Trop For Sci. 2005;17(4):574–87.
Hu P, Zhong C, Zhang Y, et al. Geographic variation in seedling morphology of Casuarina equisetifolia subsp, equisetifolia (Casuarinaceae). Aust J Bot. 2016;64(2):160–70.
Ye G, Zhang H, Chen B, et al. De novo genome assembly of the stress tolerant forest species Casuarina equisetifolia provides insight into secondary growth. Plant J. 2019;97(4):779–94.
Ren H, Xu Y, Zhao X, et al. Optimization of tissue culturing and genetic transformation protocol for Casuarina equisetifolia. Front Plant Sci. 2021;12:784566.
Ai D, Wang Y, Wei Y, et al. Comprehensive identification and expression analyses of the SnRK gene family in Casuarina equisetifolia in response to salt stress. BMC Plant Biol. 2022;22(1):572.
Wang Y, Zhang Y, Fan C, et al. Genome-wide analysis of MYB transcription factors and their responses to salt stress in Casuarina equisetifolia. BMC Plant Biol. 2021;21(1):328.
Wang Y, Zhang Y, Wei Y, et al. Characterization of HAK protein family in Casuarina equisetifolia and the positive regulatory role of CeqHAK6 and CeqHAK11 genes in response to salt tolerance. Front Plant Sci. 2022;13:1084337.
Dahro B, Wang F, Peng T, et al. PtrA/NINV, an alkaline/neutral invertase gene of Poncirus trifoliata, confers enhanced tolerance to multiple abiotic stresses by modulating ROS levels and maintaining photosynthetic efficiency. BMC Plant Biol. 2016;16:76.
Liu JH, Inoue H, Moriguchi T. Salt stress-mediated changes in free polyamine titers and expression of genes responsible for polyamine biosynthesis of apple in vitro shoots. Environ Exp Bot. 2008;62:28–35.
Thompson JD, Gibson TJ, Plewniak F, et al. The CLUSTAL_X windows interface, flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic acids res. 1997;25(24):4876–82.
Oota S. Threetree, a new method to reconstruct phylogenetic trees. Genome Inform. 1998;9:340–1.
Kumar S, Stecher G, Tamura K. MEGA7, Molecular evolutionary genetics analysis version 7,0 for bigger datasets. Mol Biol Evol. 2016;33(7):1870–4.
Chen C, Chen H, Zhang Y, et al. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020;13(8):1194–202.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402–8.
Bryfczynski S , Pargas R. GraphPad: A Graph Creation Tool for CS2/CS7[J]. SIGCSE bulletin. 2009;41(3):389.
Lescot M, Dehais P, Thijs G, et al. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002;30:325–7.
Chang W, Lee T, Huang H, et al. PlantPAN: Plant promoter analysis navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene groups. BMC Genomics. 2008;9:561.
Shannon P, Markiel A, Ozier O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13:2498–504.
Sun S, Li X, Gao S, et al. A novel WRKY transcription factor from Ipomoea trifida, ItfWRKY70, confers drought tolerance in sweet potato. Int J Mol Sci. 2022;23(2):686.
Cheng Y, Luo J, Li H, et al. Identification of the WRKY gene family and characterization of stress-responsive genes in Taraxacum kok-saghyz Rodin. Int J Mol Sci. 2022;23(18):10270.
Liu Y, Wang M, Huang Y, et al. Genome-wide identification and analysis of R2R3-MYB genes response to saline-alkali stress in Quinoa. Int J Mol Sci. 2023;24(11):9132.
Chen H, Tang K, Song H, et al. Study on 3 typical salt resistant plants for island vegetation restoration. J Oceanogr. 2017;36(03):379–84.
Wang W, Pang J, Zhang F, et al. Integrated transcriptomics and metabolomics analysis to characterize alkali stress responses in canola (Brassica napus L,). Plant Physiol Biochem. 2021;166:605–20.
Sun S, Chen H, Yang Z, et al. Identification of WRKY transcription factor family genes in Pinus massoniana Lamb, and their expression patterns and functions in response to drought stress. BMC Plant Biol. 2022;22(1):424.
Phukan UJ, Jeena GS, Shukla RK. WRKY transcription factors, molecular regulation and stress responses in plants. Front Plant Sci. 2016;7:760.
Song H, Cao Y, Zhao L, et al. Review: WRKY transcription factors: Understanding the functional divergence. Plant Sci. 2023;334:111770.
Xu H, Watanabe KA, Zhang L, et al. WRKY transcription factor genes in wild rice Oryza nivara. DNA Res. 2016;23(4):311–23.
Huang S, Gao Y, Liu J, et al. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol Genet Genomics. 2012;287(6):495–513.
Yang X, Zhou Z, Fu M, et al. Transcriptome-wide identification of WRKY family genes and their expression profiling toward salicylic acid in Camellia japonica. Plant Signal Behav. 2021;16(1):1844508.
Yin G, Xu H, **ao S, et al. The large soybean (Glycine max) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups. BMC Plant Biol. 2013;13:148–148.
Chen Y, Kong X, Yang L, et al. Genome-wide identification of WRKY family genes and the expression profiles in response to nitrogen deficiency in poplar. Genes (Basel). 2022;13(12):2324.
Wu W, Zhu S, Xu L, et al. Genome-wide identification of the Liriodendron chinense WRKY gene family and its diverse roles in response to multiple abiotic stress. BMC Plant Biol. 2022;22(1):25.
Xu Z, Sun L, Zhou Y, et al. Identification and expression analysis of the SQUAMOSA promoter-binding protein (SBP)-box gene family in Prunus mume. Mol Genet Genomics. 2015;290(5):1701–15.
Grunewald W, Smet ID, Lewis DR, et al. Transcription factor WRKY23 assists auxin distribution patterns during Arabidopsis root development through local control on flavonol biosynthesis. Proc Natl Acad Sci U S A. 2012;109(5):1554–9.
Stetter MG, Benz M, Ludewig U. Increased root hair density by loss of WRKY6 in Arabidopsis thaliana. PeerJ. 2017;5:e2891.
Ali MA, Azeem F, Nawaz MA, et al. Transcription factors WRKY11 and WRKY17 are involved in abiotic stress responses in Arabidopsis. J Plant Physiol. 2018;226:12–21.
Fan C, Qiu Z, Zeng B, et al. Physiological adaptation and gene expression analysis of Casuarina equisetifolia under salt stress. Biol Plant. 2018;62:489–500.
Wang Y, Zhang J, Qiu Z, et al. Transcriptome and structure analysis in root of Casuarina equisetifolia under NaCl treatment. PeerJ. 2021;9:e12133.
Ren H, Zhang Y, Zhong M, et al. Calcium signaling-mediated transcriptional reprogramming during abiotic stress response in plants. Theor Appl Genet. 2023;136(10):210.
Zhang M, Zhao R, Huang K, et al. The OsWRKY63-OsWRKY76-OsDREB1B module regulates chilling tolerance in rice. Plant J. 2022;112(2):383–98.
Acknowledgements
We thank Prof. Wenwu Wu and his student Liangyu Guo for providing the assistance in transcriptome analysis.
Funding
This work was supported by the National Key R&D Program for Young Scientists (grant number: 2021YFD2200900), the State Key Laboratory of Subtropical Silviculture (grant number: SKLSS-KF2022-08).
Author information
Authors and Affiliations
Contributions
XHZ performed the genome-wide analysis of CeqWRKYs. XHZ and GNQ conducted the experiments, and wrote the manuscript. JHL assisted with the experiments. KC assisted with the data analysis. XXM cultured the plant materials. JH revised the manuscript. SKL and HMR were responsible for the overall concept and experimental designs, analysis, and manuscript preparation. All authors approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhao, X., Qi, G., Liu, J. et al. Genome-wide identification of WRKY transcription factors in Casuarina equisetifolia and the function analysis of CeqWRKY11 in response to NaCl/NaHCO3 stresses. BMC Plant Biol 24, 376 (2024). https://doi.org/10.1186/s12870-024-04889-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-024-04889-w