
Abstract
Intercrop** can obtain yield advantages, but the mechanism of yield advantages of maize-legume intercrop** is still unclear. Then, we explored the effects of crop** systems and N input on yield advantages in a two-year experiment. Crop** systems included monoculture maize (Zea mays L.) (MM), monoculture soybean (Glycine max L. Merr.) (MS), monoculture peanut (Arachis hypogaea L.) (MP), maize-soybean substitutive relay intercrop** (IMS), and maize-peanut substitutive strip intercrop** (IMP). N input included without N (N0) and N addition (N1). Results showed that maize’s leaf area index was 31.0% and 34.6% higher in IMS and IMP than in MM. The specific leaf weight and chlorophyll a (chl a) of maize were notably higher by 8.0% and 18.8% in IMS, 3.1%, and 18.6% in IMP compared with MM. Finally, N addition resulted in a higher thousand kernels weight of maize in IMS and IMP than that in MM. More dry matter accumulated and partitioned to the grain, maize's averaged partial land equivalent ratio and the net effect were 0.76 and 2.75 t ha−1 in IMS, 0.78 and 2.83 t ha−1 in IMP. The leaf area index and specific leaf weight of intercropped soybean were 16.8% and 26% higher than MS. Although soybean suffers from shade during coexistence, recovered growth strengthens leaf functional traits and increases dry matter accumulation. The averaged partial land equivalent ratio and the net effect of intercropped soybean were 0.76 and 0.47 t ha−1. The leaf area index and specific leaf weight of peanuts in IMP were 69.1% and 14.4% lower than in the MP. The chlorophyll a and chlorophyll b of peanut in MP were 17.0% and 24.4% higher than in IMP. A less dry matter was partitioned to the grain for intercropped peanut. The averaged pLER and NE of intercropped peanuts were 0.26 and -0.55 t ha−1. In conclusion, the strengthened leaf functional traits promote dry matter accumulation, maize-soybean relay intercrop** obtained a win–win yield advantage, and maize-peanut strip intercrop** achieved a trade-off yield advantage.
Similar content being viewed by others


Introduction
Agriculture production faces great challenges, such as increasing farmland productivity with limited resources to feed the continuously growing global population [1]. Intercrop**, defined as two or more crops grown in the same land for the whole or a part of their growing period [2], has been used worldwide because of its high farmland productivity [3], efficient use of nutrients [4], better control of diseases and pests [2, 5], and its possibility for providing a sustainable agricultural development [6].
The yield advantages of intercrop** systems are mainly based on the complementary or competitive use of resources [7]. However, the yield advantage of maize varies when intercropped with different legumes [3]. Although maize-soybean and maize-peanut intercrop** achieves a land equivalent ratio (LER) of higher than one, maize yield increased while soybean and peanut yield decreased [7]. Reasonable N input in intercrop** will improve the grain yield advantages [8]. For instance, the maize-legume intercrop** benefits more from the complementary effect than the selection effect without N addition, but N fertilization led to a more robust yield advantage by shifting the complementary effect to the selection effect [7, 9, 10]. Although intercrop** cereal with legumes can benefit cereal by strengthening the symbiosis nitrogen fixation of legumes and increasing soil N pool [11, 12], N input and crop** systems affected the N fixation ability and yield advantage of legumes in the intercrop** [8, 13]. High N input inhibits the symbiosis of nitrogen fixation and decreases the grain yield of legumes, but the competitive use of N by cereal can alleviate the inhibiting effect [14]. Compared with wheat-faba bean intercrop**, the grain yield and biological N fixation of faba bean improved in maize-faba bean intercrop** [15]. To date, some studies have revealed the yield advantages mechanism of maize-legume intercrop** from the standpoint of ecology [7, 16]. However, the yield advantages mechanism of maize-legume intercrop** in physiology is still poorly understood, especially from leaf functions to yield formation and yield advantages in intercrop**.
Biomass accumulation is the basis of yield, and the crop leaf functional traits affect biomass production. The leaf functional traits, e.g., leaf area (LA), leaf area index (LAI), specific leaf weight (SLW), chlorophyll content, and chloroplast ultrastructure, are used to assess the characteristics of crop growth on a temporal scale [17,18,19]. On the temporal and spatial scales, relative growth rates (RGR) help evaluate crop growth in different periods [20]. Besides, the crop growth environment plays a crucial role in photosynthesis, such as shading affecting the leaf morphology, structure, function, and biomass accumulation [21, 22]. Compared with normal light, shading increases soybean leaf chloroplast number, grana lamella thickness, and photosynthetic pigment per unit mass while decreasing the net photosynthetic rate and the chloroplast and starch grain size [18]. After maize harvest, the full sunshine condition is beneficial to increase the aboveground biomass, leaf thickness, and chlorophyll content of the intercropped soybean [23]. The recovery growth promotes biomass accumulation and improves soybean grain yield [19].
Previous studies have focused on ecology and agronomy aspects to investigate the yield advantage of intercrop** [7, 16, 24], but few studies evaluate the leaf functional traits and dry matter partition on the yield advantage in maize-legume intercrop**. Therefore, the objectives of this study were (1) to investigate the effects of crop** systems and N addition on crop leaf functional traits, e.g., leaf area index, specific leaf weight, and chloroplast ultrastructure, (2) to analyze the effect of crop** systems and N addition on crops dry matter accumulation and partition (3) to reveal relationships between the crop leaf functional traits, dry matter accumulation and partition, and yield advantage in maize-legume intercrop**.
Material and methods
Site description, experimental design and sampling
The field experiment was conducted in Renshou County in 2017–2018 (29°60’ N, 104°00’ E), Sichuan Province, Southwest China. The climate of the field region was subtropical monsoon humid, with an average annual temperature of 17.4 °C, rainfall of 1009.4 mm, and sunshine of 1196.6 h. Before planting, soil fertility was investigated. The soil pH, organic matter, total N, total P, and total K were 8.18, 14.19 g kg−1, 1.22 g kg−1, 1.95 g kg−1, and 26.06 g kg−1. The maize (** systems. The N input levels included N addition (N1) and without N (N0). In N addition treatments, urea was used as the N fertilizer, and the rate of fertilizer used in the current study was according to the local production requirement. The N fertilizer for monoculture crop** was 80 kg N ha−1 for legumes and 240 kg N ha−1 for maize. The N fertilizer for the intercrop** was half of the monoculture crop** because a replacement intercrop** with a replace ratio of 0.5 was used. Namely, the N fertilizer of intercrop** system was 160 kg N ha−1, including 40 kg N ha−1 for legumes and 120 kg N ha−1 for maize (Table 1). The per-plant fertilizer was the same for the same crop in intercrop** and monoculture.
In the N addition treatment, the N fertilizer for intercrop** and monoculture maize was divided into base fertilizer (80 kg N ha−1) and topdressing (the rest of the N fertilizer). The topdressing was separated into two equal parts and applied at the V6 and V12 stages of maize (Table 1). In intercrop**, the legumes base fertilizer (40 kg N ha−1) was mixed and applied with maize topdressing at the V12 stage of maize. In the N addition treatment of monoculture legumes, the N fertilizer for monoculture soybean and peanut was used as the base fertilizer at 80 kg ha−1. The P (calcium superphosphate) and K (potassium chloride) fertilizers were used as base fertilizers at 120 kg P2O5 ha−1 and 100 kg K2O ha−1 for crops. The base fertilizer was strip placement 5 cm away from the seeds when sowing crops. The topdressing of maize was strip placement 5 cm away from the plants.
Crop** systems included monoculture maize (MM), monoculture soybean (MS), monoculture peanut (MP), maize-soybean substitutive relay intercrop** (IMS), and maize-peanut substitutive strip intercrop** (IMP) (Fig. 1), with 2:2 row proportions under replacement series. The component species are almost simultaneous sowing in the strip intercrop** systems, but the maturing species is interplanted with seeds of the following species in relay intercrop** systems [25]. Three replicates for each treatment resulted in a total of thirty plots. The plot size was 6.0 m (width) × 5.8 m (length). An equal row spacing (0.5 m) planting method was adopted for all the crop** systems (Fig. 1). In all crop rows, one seedling per hole. The plant spacing was 0.2 m, and the density of maize was 100,000 plants ha−1 for M and 50,000 plants ha−1 for IMS and IMP. Maize was manually sown on April 8, 2017, and April 4, 2018, and harvested on August 4, 2017, and July 31, 2018. The plant spacing was 0.1 m, and the density of monoculture legumes (MS or MP) was 200,000 plants ha−1 and 100,000 plants ha−1 for intercropped legumes. Soybean was manually sown on June 10, 2017, and June 4, 2018, and harvested on November 1, 2017, and November 3, 2018. Peanut was manually sown on April 10, 2017, and April 9, 2018, and harvested on August 10, 2017, and August 31, 2018.

Schematic diagram of crop** system. Monoculture maize (MM), monoculture soybean (MS), monoculture peanut (MP), maize-soybean relay intercrop** (IMS), maize-peanut strip intercrop** (IMP)
Sampling and measurements
Leaf area index and specific leaf weight
Three individual plants from each plot were collected at the silking stage (R1) and milk stage (R3) of maize, the fifth trifoliolate stage (V5) and the full pod stage (R4) of soybean, and the beginning bloom stage (R1) and the full seed stage (R6) of peanut in the middle section of the middle row.
Maize leaf area was calculated as length multiplied by width and coefficient K, and the K is 0.75 [19]. Four leaf discs (diameter = 1.2 cm) were punched from each of the third leaves from the top of the soybean, and ten leaves were sampled. One leaf disc (diameter = 1.2 cm) was punched for each of the third leaves from the top of the top in peanut, and twenty leaves were sampled. The leaf discs and the sample leaf were over-dried at 75 °C to constant. Then the dry weight of the leaf was determined, and calculate leaf area. The leaf area was calculated as follows:
where LA, LAdisc, DMdisc, and DMtotal are the leaf area, leaf area of the discs, dry matter of the discs, and total leaf dry weight.
The observed and expected leaf area index (LAI) [26] was calculated as follows:
where LAIobs, LAobs_A, LAobs_B, and Land area are the observed leaf area index of the intercrop** system, the observed leaf area of species A, the observed leaf area of species B, and the land area occupied by crops.
where LAIexp, LAexp_A, LAexp_B, and Land area are the expected leaf area index of the intercrop** system, the expected leaf area of species A, the expected leaf area of species B, and the land area occupied by crops.
where LAmono_A, LAmono_B, pA, and pB are the leaf area of monoculture species A, monoculture species B, the land sharing ratio of species A, and the land sharing ratio of species B.
The specific leaf weight (SLW) [27] was calculated as follows:
where DMtotal and LA are the total leaf dry weight and area.
Chlorophyll and chloroplast ultrastructure
Leaf samples of three individual plants from each plot were collected at the R1 and R3 stages of maize (ear leaf) [28], at the V5 and R4 stages of soybean (the third leaf from the top) [29, 30], and at the R1 and R6 stages of peanuts (the third leaf from the top) [31]. The leaf chlorophyll was extracted and determined using the method of Arnon [32]. Samples for chloroplast ultrastructure observation were fixed with a mixture of 3% glutaraldehyde and embedded to make semi-thin sections. The semi-thin sections were optically positioned, and ultrathin sections were stained with uranyl acetate and lead citrate, then examined with a transmission electron microscope (TEM; HITACHI, H-600IV, Japan) [18].
Measurements of dry matter and yield
Plant samples were collected at the V12, R1, R3, and R6 stages of maize, the V5, R2, R4, R6, and R8 stages of soybean, and the R1, R2, R4, and R6 stages of peanut. Three plants were sampled from each plot. These plant samples were separated into different organs: stem, leaf, and grain. Then, samples were oven-dried to constant weight at 75 °C and weighed.
Relative Growth Rate (RGR) is defined as growth in terms of a rate of increase in size per unit of size. The mean RGR over an interval of time between t1 and t2 is usually calculated as shown in the following formula [33]:
where W2 and W1 are the dry matter at the sampling dates two and one, and t2 and t1 are the sampling dates, respectively.
At the mature stage, twenty consecutive plants of maize, soybean, and peanut were harvested in the middle row of each plot, and yield components were investigated. The net effect (NE) was used to assess the yield advantage of intercrop** [7, 34], and the NE of two species of intercrop** systems was calculated as follows [35]:
where NE, NE1, and NE2 are the net effect of intercrop**, species one, and species two.
where NEi is the net effect of intercrop** species i, Yobs_i, and Yexp_i are the observed and expected yields of species i (tonne per ha).
where Mi and pi are the monoculture crop** yield (tonne per ha) and farmland sharing ratio of species i (%).
The land equivalent ratio (LER) was used to determine the land use advantage in intercrop** and calculated as follows [36]:
where pLERm and pLERL are the partial land equivalent ratio of maize and legumes. Yi_m and Ys_m are maize grain yields in intercrop** and monoculture crop**. Yi_L and Ys_L are legumes grain yields in intercrop** and monoculture crop**.
Statistical and analysis
All statistical analyses were conducted with R v 4.4.2 [37]. We conducted a one-way ANOVA to test the effects of the crop** system, N levels, and growing seasons on crops’ LAI, SLW, RGR, biomass, and yield components, as well as on the pLER and NE of the component crops and the intercrop** system. The one-way ANOVA (Tukey HSD, p < 0.05) was performed with HSD.test() functions in the R package agricolae v 1.3–6 [38]. Figures were drawn by GraphPad Prism 9 (GraphPad Software, San Diego, California, USA).
Results
Leaf Area Index (LAI) and Specific Leaf Weight (SLW)
Nitrogen addition considerably enhanced maize’s LAI in 2018, but the LAI was independent of N addition in 2017 (Fig. 2A). The LAI of maize was 31.0% and 34.6% higher in IMS and IMP than in MM (2.26, mean of two years) (Fig. 2A). The LAI of soybean was independent of N addition at the V5 stage, but at the R4 stage, it was notably increased with N addition (Fig. 2B). The LAI of soybean in IMS (0.36, mean of two years) was 11.5% lower than in MS at the V5 stage. At the R4 stage, the leaf area index of intercropped soybean was 16.8% higher than that in MS in 2017, while that of intercropped soybean was lower than MS in 2018 (Fig. 2B). Similarly, the LAI of peanut was independent of N addition. Crop** seasons affected peanuts’ LAI. At the R1 stage, the LAI of peanuts was remarkably higher in 2018 than in 2017. In contrast, the LAI of peanut was greater in 2017 than in 2018 at the R4 stage (Fig. 2C). Moreover, the LAI of monoculture peanut (1.28, mean of two years) was 50.1% greater than that in IMP at the R6 stage.

Effects of nitrogen input and crop** system on leaf area index (LAI). MM, monoculture maize, MS, monoculture soybean, MP, monoculture peanut, IMS, maize-soybean relay intercrop**, IMP, maize-peanut strip intercrop**. Panels A (maize), R1, the silking stage, and R3, the milk stage. Panels B (soybean), V5, the fifth trifoliolate stage, and R4, the full pod stage. Panels C (peanut), R1, the beginning bloom stage, and R6, the full seed stage. N0, 0 kg N ha−1, N1, 80 kg N ha.−1. Data were shown as mean with S.D. Different lower case letter donates significant difference between crop** systems under the same N input (Tukey HSD, p < 0.05). The Tukey HSD values were shown as standard bar above each N treatments. Results of the one-way ANOVA were displayed at the top of each panel, N, N input, C, crop** system, and ‘*’ and ‘ns’ represent significant and insignificant difference at the same growth stage (Tukey HSD, p < 0.05)
N addition enhanced maize’s SLW in 2018, but it was insignificant in 2017 (Fig. S2A). The SLW of maize in IMS and IMP was 8% and 3.1% higher than that of MM (54.3 g m−2, mean of two years). Compared with MS, the SLW of intercropped soybean (48.3 and 50.7 g m−2, mean of two years) was notably higher by 7.9% under N0 and 7.1% under N1 at the R4 stage (Fig. S2B). Regarding peanut, intercrop** significantly increased peanuts’ SLW in 2018. N addition remarkably enhanced SLW but intercrop** resulted in a notable decrease in SLW at the R1 stage (Fig. S2C). The SLW of MP was 54.6% higher than that of IMP (21.84 g m−2, mean of two years) at the R6 stage.
Chloroplast ultrastructure and chlorophyll (Chl) content
Nitrogen addition strengthened crop leaf physiological functional traits, such as promoting the development of chloroplast, increasing the number of chloroplast grana, and enhancing the accumulation of starch grana in the chloroplast (Figs. 3–5). The grana lamellae of maize leaves in IMS and IMP were thickened in MM (Fig. 3). In contrast to MM, the chloroplast volume of maize leaves was greater in IMS and IMP, and it was bigger in IMS than in IMP. In monoculture, the thylakoid of soybean leaves was elongated, and grana stacking was less at the V5 stage. In contrast, the thylakoid of soybean leaves in IMS was round and closely arranged with more starch grains, which occupied as much as 60% of the chloroplast (Fig. 4). Similarly, chloroplast volume and grana lamellae of soybean leaves were greater in IMS than in the monoculture at the R4 stage. The chloroplast volume of peanut leaves was lower in IMP than in the monoculture; however, more and larger grana lamellae and starch grains were observed in IMP than in the monoculture (Fig. 5). Similarly, the starch grains accounting for more than 60% of the chloroplast in peanut leaves (Fig. 5).

Effects of nitrogen input and crop** system on the chloroplast ultrastructure of maize functional leaf in 2018. R1, MM, monoculture maize, IMS, maize-soybean relay intercrop**, and IMP, maize-peanut strip intercrop**. N0, 0 kg N ha−1, and N1, 80 kg N ha−1. R1, the silking stage, and R3, the milk stage. Mt, Mitochondria, Gl, Grana lamella, Sl, Stroma lamella, Sg, Starch grain, Og, Osmiophilic granules. Panels A-F, Chloroplast ultrastructure of maize at the silking stage (R1), and panels G-L, Chloroplast ultrastructure of maize at the milk stage

Effects of nitrogen input and crop** system on the chloroplast ultrastructure of soybean functional leaf in 2018. MS, monoculture soybean, and IMS, soybean in the maize-soybean relay intercrop**. N0, 0 kg N ha−1, and N1, 80 kg N ha−1. V5, the fifth trifoliolate stage, and R4, the full pod stage. Mt, Mitochondria, Gl, Grana lamella, Sl, Stroma lamella, Sg, Starch grain, Og, Osmiophilic granules. Panels A-D, Chloroplast ultrastructure of soybean at the fifth trifoliolate stage, and panels E–H, Chloroplast ultrastructure of soybean the full pod stage

Effects of nitrogen input and crop** system on the chloroplast ultrastructure of peanut functional leaf in 2018. MP, monoculture peanut, and IMP, peanut in maize-peanut strip intercrop**. N0, 0 kg N ha−1, and N1, 80 kg N ha−1. R1, the beginning bloom stage, and R6, the full seed stage. Mt, Mitochondria, Gl, Grana lamella, Sl, Stroma lamella, Sg, Starch grain, Og, Osmiophilic granules. Panels A-D, Chloroplast ultrastructure of peanut at the beginning bloom stage, and panels E–H, Chloroplast ultrastructure of soybean at the full seed stage
Generally, nitrogen addition and intercrop** notably affected crop leaf Chl a, Chl b, and Chl a/b (Fig. 6). The Chl a of maize in IMS and IMP was 18.8% and 18.6% higher than that in MM (2.72 mg g−1), while the Chl b was higher by 16.3% in IMS and 18.2% in IMP compared with MM (0.86 mg g−1), respectively (Fig. 6A). Although N addition decreased the Chl a/b ratio of maize, intercrop** maize with legumes and N addition improved the Chl a/b ratio (Fig. 6A).

Effects of nitrogen input and crop** system on leaf chlorophyll content (2018). MM, monoculture maize, MS, monoculture soybean, MP, monoculture peanut, IMS, maize-soybean relay intercrop**, IMP, maize-peanut strip intercrop**. Chl a, Chlorophyll a, Chl b, Chlorophyll b, Chl a/b, Chlorophyll a/b ratio. Panels A (maize), R1, the silking stage, and R3, the milk stage. Panels B (soybean), V5, the fifth trifoliolate stage, and R4, the full pod stage. Panels C (peanut), R1, the beginning bloom stage, and R6, the full seed stage. N0, 0 kg N ha−1, N1, 80 kg N ha.−1. Data were shown as mean with S.D. Different lower case letter donates significant difference between crop** systems under the same N input (Tukey HSD, p < 0.05). The Tukey HSD values were shown as standard bar above each N treatments. Results of the one-way ANOVA were displayed at the top of each panel, N, N input, C, crop** system, and ‘*’ and ‘ns’ represent significant and insignificant difference at the same growth stage (Tukey HSD, p < 0.05)
Regarding soybean (Fig. 6B), the Chl b content of soybean leaves in IMS at the R4 stage was 13.9% lower than that in the monoculture. The Chl a of soybean leaves in monoculture was 6.9% higher than in IMS with N addition (2.19 mg g−1). Independent of N addition, the Chl a/b ratio of soybean leaves was lower in IMS than in MS at the V5 stage, in contrast, the Chl a/b ratio was greater in intercropped soybean than in MS at the R4 stage (Fig. 6B). Focusing on peanuts (Fig. 6C), the contents of Chl a, Chl b, and Chl a/b decreased with the growth. The Chl a and Chl b of peanut leaves in IMP were 16.0% and 22.8% higher than the monoculture (1.45 and 0.47 mg g−1) with N addition. Besides, intercrop** decreased the Chl a/b ratio of peanuts leaves at the R6 stage.
Dry matter accumulation and partition
The crop** season affected maize per-plant dry matter accumulation, except for the V12 stage, maize biomass was significantly higher in 2018 than in 2017. Except for the V12 stage, nitrogen addition notably affected maize per-plant dry matter accumulation (Fig. 7A). At the R6 stage, the per plant dry matter accumulation of maize in MM, IMS, and IMP increased by 15.3%, 20.0%, and 15.6% in N1 compared with N0, respectively. The dry matter accumulation of maize per plant in IMS and IMP 44.7% and 53.0% higher than that in MM (23.89 g plant−1, mean of two years), respectively. In zero N treatment, the biomass of MM was lower in 2018 than in 2017. With N addition, the biomass of intercropped maize was higher in 2018 than in 2017. (Figs. 7A and S3A, B).The accumulated dry matter of maize was mainly transported to the ear, and the dry matter of maize ear was greater in IMS and IMP than MM (Figs. 7A and S3A, B). With N addition, the RGR of maize was higher in both IMS and IMP than in MM at the R3-R6 period (Fig. 8A).

Effects of nitrogen input and crop** system on dry matter accumulation. MM, monoculture maize, MS, monoculture soybean, MP, monoculture peanut, IMS, maize-soybean relay intercrop**, IMP, maize-peanut strip intercrop**. Panels A (maize), V12, the twelfth leaf stage, R1, the silking stage, R3, the milk stage, R6, the maturity stage. Panels B (soybean), V5, the fifth trifoliolate stage, R2, the full bloom stage, R4, the full pod stage, R6, the full seed stage, and R8, maturity stage. Panels C (peanut), R1, the beginning bloom stage, R2, the beginning ped stage, R4, the full pod stage, R6, the full seed stage. N0, 0 kg N ha−1, N1, 80 kg N ha−1. Data were shown as mean with S.D. Different lower case letter donates significant difference between crop** systems under the same N input (Tukey HSD, p < 0.05). The Tukey HSD values were shown as standard bar above each N treatments. Results of the one-way ANOVA were displayed at the top of each panel, N, N input, C, crop** system, and ‘*’ and ‘ns’ represent significant and insignificant difference at the same growth stage (Tukey HSD, p < 0.05)

Effects of nitrogen input and crop** system on crops’ relative growth rate. MM, monoculture maize, MS, monoculture soybean, MP, monoculture peanut, IMS, maize-soybean relay intercrop**, IMP, maize-peanut strip intercrop**. Panels A (maize), V12-R1, from the twelfth leaf stage to silking stage, R1-R3, from the silking stage to milk stage, and R3-R6, from the milk stage to maturity stage. Panels B (soybean), V5-R2, from the fifth trifoliolate stage to full bloom stage, R2-R4, from the full bloom stage to full pod stage, and R4-R6, from the full pod stage to full seed stage. Panels C (peanut), R1-R2, the beginning bloom stage to beginning ped stage, R2-R4, from the beginning pod stage to full pod stage, R4-R6, from the full pod stage to full seed stage. N0, 0 kg N ha−1, N1, 80 kg N ha.−1. Data were shown as mean with S.D. Different lower case letter donates significant difference between crop** systems under the same N input (Tukey HSD, p < 0.05). The Tukey HSD values were shown as standard bar above each N treatments. Results of the one-way ANOVA were displayed at the top of each panel, N, N input, C, crop** system, and ‘*’ and ‘ns’ represent significant and insignificant difference at the same growth stage (Tukey HSD, p < 0.05)
The biomass of soybean was significantly lower in 2018 than in 2017. Although soybean per plant dry matter was independent of N input at the V5 stage, N addition notably enhanced soybean dry matter accumulation at the reproductive growth stage (Fig. 7B). Compared with without N, soybean dry matter accumulation of MS (68.3 g plant−1, mean of two years) and IMS (79.6 g plant−1) was 16.5% and 12.5% higher in N1 at the R8 stage. The dry matter partitioned to the grain of soybean in IMS enhanced by N addition treatment, namely by 6.2% and 8.5% at the R4 and R6 stages, respectively (Fig. S3C, D). Meanwhile, the RGR of soybean was higher in 2018 than in 2017 at the V5-R2 and R2-R4 periods. The RGR of intercropped soybean was 4.9% and 5.5% higher in N1 than in N0 at the V5-R2 and R2-R4 periods (Fig. 8B). Finally, the average grain yield of intercropped soybean was 13.8% higher in N1 than in N0. The accumulation of soybean per plant dry matter was 35.5% lower in IMS than in MS (2.58 g plant−1, mean of two years) at the V5 stage (Fig. 7B). With soybean growth, a more robust increase of dry matter accumulation was obtained in IMS than in MS. The dry matter accumulation of soybean was 28.4% lower at the V5 stage and 14.4% higher in IMS at the R8 stage compared with MS (two-year average). Compared with MS, the dry matter allocation to soybean stem in IMS decreased, while dry matter allocation to soybean leaf and pod increased, especially in R6 and R8 (Fig. S3C, D). The RGR of soybean was 17.6% greater in MS than in IMS at the V5-R2 period; in contrast, it was 9.9% higher in IMS than in MS at the R2-R4 period (Fig. 8B).
The peanut per-plant dry matter accumulation was higher in 2018 than in 2017 at the R1 and R2 stages. The averaged per-plant dry matter of intercropped peanut (27 g plant−1) was 30% higher in N1 than in N0 at the R6 stage (Fig. 7C). More dry matter was partitioned to grain in intercropped peanut than in MP at the harvest stage (Fig. S3E, F). The RGR of intercropped peanut with N addition was 30% higher than without N (Fig. 8C). Intercrop** harmed the dry matter accumulation of peanuts (Fig. 7C). Compared with MP (46.1 g plant−1), dry matter accumulation of peanuts was 48.1% lower in IMP in two years on average at the R6 stage. At the R1 stage, the dry matter allocation of peanuts to stems was fewer but more to leaves. As peanuts grew, dry matter partitioned to the pod gradually increased, but the partition to stem and leaf decreased (Fig. S3E, F). At maturity, dry matter partitioned to stem and pod gradually increased with the peanut growing in the monoculture, but less dry matter was partitioned to the pod in intercropped peanut. The RGR of peanuts (0.11 g day−1) was 58.1% significantly higher in MP than in IMP at the R1-R2 stage (Fig. 8C).
Yield advantages of intercrop**
Intercrop** with legumes significantly increased maize kernel number and thousand kernels weight, but the magnitude of that increase depended on crop** seasons. In contrast, the kernel number of monoculture maize reduced with two years of zero N input (Table 2). In addition, maize kernel number and weight remarkably improved in 2017 and 2018 (Table 2). The per-plant kernel number of maize peaked in IMP (478.68 kernels), and the thousand kernels weight of maize in IMS (349.94 g) was the highest. Regarding soybean, the number of per-plant grains was notably 25% higher in IMS than in MS (98.89 seeds), and the number of per-plant grains was notably 13.5% higher in N addition compared with no N (104.25 g plant−1) (Table 2). The hundred-seed weight of soybean was improved when intercropped with maize or N addition. Generally, the number of per-plant peanuts grains was independent in N addition, but it was remarkably 49.9% lower in IMP than in MP (16.62 seeds). A lower hundred-seed weight of peanut was obtained when intercropped with maize, but it increased by 5.6% with N addition (60.66 g) (Table 2). In summary, intercropped maize with N addition obtained more kernels and greater seeds size; in contrast, the better growth of maize had disadvantageous effects on legumes growth and led to fewer seeds and smaller seeds size (Table 2).
Generally, Maize in IMP showed a higher pLER and net effect than IMS, but there were no differences between crop** systems. Compared with without N, N addition decreased pLER by 0.91% and enhanced net effect by 10.2% for maize in IMP (pLERM, 0.75, and net effect, 2.94 t ha−1 mean of two years) (Table 3). On average for two-year, soybean intercropped with maize showed a pLER and net effect of 0.75 and 0.44 t ha−1 in N0 and by 0.78 and 0.50 t ha−1 in N1 (Table 3). Although N addition led to a decrease in pLER by 0.1%, the net effect of soybean was 14.9% higher with N addition than without N (0.435 t ha−1) at two years average (Table 3). Although the pLER of peanut increased by N addition, the net effect of peanut decreased by N addition (Table 3). Finally, the LER (1.52) and net effect (3.21 t ha−1) of IMS were 25% and 40.6% higher than those of IMP (Table 3). In IMS, the LER and the net effect were 4.02% and 19.3% higher in N addition than without N (1.49 t ha−1).
Discussion
Performances of maize in maize-soybean and maize-peanut substitutive intercrop**
Due to the taller plant height of maize, maize dominates and shades legumes in the maize-legume intercrop** systems, intercrop** with legumes and N addition was beneficial to increasing the LAI and SLW of maize (Figs. 2 and S2). Soybean was sown at the V12 stage of maize, and maize was harvested at the R1 stage of soybean. Leaf trait plays a critical role in light use and biomass accumulation, such as LAI and SLW directly affect light intercept and photosynthesis [39]. A higher LAI denotes more leaf area on the unit farmland and greater light interception by crop. Meanwhile, a greater SLW indicates leaves are thicker. Amanullah [40] pointed out that high-yield cultivars obtain thicker leaves and higher photosynthetic capacities. Indeed, consistent results were observed that thicker leaves were associated with a more robust synthesis of starch in leaves. The maize leaf chloroplast volume improved and grana lamellae stacking thickened when intercropped with legumes and N addition (Fig. 3).
A lower N condition reduces leaf chlorophyll content, and thinner leaves usually obtain lower chlorophyll content [40]. In the current study, the chlorophyll of maize was higher in IMS and IMP than in monoculture maize (Fig. 6). But the leaf functional traits of maize in IMP were more robust than in IMS (Figs. 3 and 6). On the one hand, light is absorbed by chlorophyll and transformed into organic compounds in the chloroplast [41]. The photoreaction is localized in the internal chloroplast membrane, called thylakoid; thus, the structure and quantity of thylakoids are decisive in effective photosynthesis [42,43,44]. As light irradiance reduced, the numbers of chloroplasts and grana lamellae increased, while the shape of chloroplast changed from ellipse or olive to swollen oblate or spheroidal. On the other hand, the Chl a content has also been suggested as one of the most decisive factors; namely, increasing Chl a can increase the photosynthetic rate [45, 46]. When intercropped with legumes, maize enhanced the light capture and was used through increasing LAI and SLW, enhancing leaf chlorophyll content, and strengthening leaf photosynthetic product accumulation. Therefore, the strengthened light-use ability is the base of biomass accumulation and yield.
Indeed, the improved leaf traits enhanced maize’s relative growth ratio in intercrop** systems (Fig. 9). Because of that, intercropped maize obtained a greater dry matter accumulation and grain yield than monoculture maize. Probably due to legume height being lower than maize, maize intercepted more light in intercrop** than monoculture [47]. Although the belowground interactions increase maize biomass and grain yield in maize-peanut substitutive strip intercrop** [48], the belowground interactions are independent of maize yield in the additive relay strip intercrop** [49]. The different maize responses to the below ground are probably due to the wide interspecific distance in the maize-soybean intercrop** (60 cm) than the maize-peanut intercrop** (35 cm) [48, 49]. Wangiyana et al. [50] also found that leaves of red rice at anthesis much greener (indicating higher N) in intercrop** with peanut resulting in much higher grain yield than in monocropped rice. Moreover, Nitrogen addition further enhances intercrop** maize’s dry matter accumulation and grain yield. Intercrop** sweet corn with peanut, presumably increasing N supply to sweet corn, was also reported to increase number of green leaves, biomass and cob weight of sweet corn per plant [51].

Model diagram of N inputs and crop** system on crops’ leaf functional traits and dry matter partition. Data of the partial land equivalent ratio (pLER) and net effect (NE) were shown as values (mean of two year) of the component crops under different N inputs. Data of the seed partition at the maturity stage, leaf area index (LAI), specific leaf weight (SLW), and relative growth rates (RGR) were shown as the relative changes (mean of two year) in intercrop** compared with the corresponding monoculture crop**. The subscript denotes the corresponding growth stages of the crop
The kernel per ear and thousand kernels weight of maize increased when intercropped with legumes (Table 2). Although the kernel per ear of maize was higher in IMP than in IMS, the thousand kernels weight of maize was greater in IMS than in IMP. Sufficient rainfall facilitated acquisition of soil nutrients, while excessive rainfall at the grain filling stage (July 2018) also means a limitation on maize grains filling (Fig. S1). Therefore the kernel number of maize was greater in 2018 than in 2017, but the kernels weight of maize was lower in 2018 than in 2017 (Table 2). Besides, N addition notably increase the yield components of intercropped maize. Long-term intercrop** enhances soil fertility and crop grain yield [4]. The improved nutrient condition is the belowground interaction mechanism of high-yielding of intercrop**. Legumes can fix nitrogen and increase soil N pool, and peanut facilitates maize Fe and Zn nutrition in intercrop** systems [52]. Therefore, interspecific facilitation improves crop nutritional status and promotes crop growth [52]. The consistent results were observed that the net effect and pLER of maize without N were 2.5 t ha−1 and 0.75 when intercropped with soybean, and N addition resulted in a greater net effect and pLER by 3.0 t ha−1 and 0.76 (Fig. 9). Although N addition led to a slight decrease in pLER of maize when intercropped with peanut, the NE of maize shift from 2.6 to 2.9 t ha−1 (Table 3).
Physiological adjustment of the intercropped legumes under different shade times
The shorter component species suffer from the shading of the taller species in intercrop** systems [19, 53]. Although the competitive use of light adversely affected the LAI and SLW of legumes during the coexistence period, the LAI and SLW of soybean can be recovered after the maize harvest (Figs. 2 and S2). In contrast, the penalty on peanuts’ LAI and SLW were not alleviated after the maize harvest (Figs. 2 and S2). The duration and intensity of shade in soybean and peanut differed, leading to a small peanut leaf area and a weak leaf functional trait. Therefore, soybean recovered after the coexistence and somewhat improved the leaf functional traits. The variation of legumes’ LAI and SLW was mainly caused by the shift of light environment in the intercrop** systems. Shading decreases the LAI and SLW of intercropped soybean by reducing leaf size and thickness in the maize-soybean intercrop** [18, 54]. After the maize harvest, the compensatory growth of soybean can eliminate the yield disadvantage [19]. Interestingly, although the recovered growth of soybean increase LAI and SLW, it was slightly different during the two crop** seasons. This was probably due to the less precipitation in 2017 than in 2018 (Fig. S1), which limited maize growth (Fig. 2A). Water deficiency will limit maize LAI and decrease grain yield [13]; in contrast, an alleviated competitive use of resources between maize and soybean results in a yield advantage of soybean [55].
The total chlorophyll of soybean significantly decreased when intercropped with maize, while the total chlorophyll of soybean was notably enhanced by N addition. The inference of crop** systems and N inputs on the chlorophyll mainly resulted from Chl a rather than Chl b (Fig. 6B). A higher chlorophyll contributes to utilizing light more efficiently, especially in a shaded environment [18]. Plants growing in the shade can optimize their light absorption efficiency by increasing pigment density per unit leaf area [56]. The increase in Chl b is most likely due to changes in the light-harvesting organization [57].
In contrast, intercrop** and N addition notably enhanced peanuts’ Chl a and Chl b (Fig. 6C). In other words, peanuts suffer from a heavier shading than soybean when intercropped with maize, attributed to a relatively extended coexistence period (Fig. 1F). This adjustment reduced the respiratory demand to help compensate for the significantly decreased photosynthetic capacity of leaves [58]. Moreover, the Chl a/b of intercropped maize decreased through the quick increase in Chl a; N addition can alleviate the penalty on Chl a/b (Fig. 6A). The Chl a/b of the legume was reduced during the coexistence period through a quick increase in Chl b. The previous study indicates that the enhancement of Chl b is beneficial to increase the light harvest pigment complex, then improve the adaptability of plants to the shade condition [59].
Transmission electron microscopy showed that N addition could increase the chloroplast volume and thicken the grana lamellae of soybean and peanut (Figs. 4–5). At the V5 stage, the thylakoids of monoculture soybean were long, and grana lamellae decreased; in contrast, the thylakoids of intercropped soybean were round and starch grains closely arranged (Fig. 4). At the R4 stage, the chloroplast volume and grana lamellae of intercropped soybean increased while the arrangement was clear. Although the chloroplast volume of intercropped peanut decreased compared with monoculture peanut, the grana lamellae and starch grains of intercropped peanut increased (Fig. 5). The shading of maize led to more grana containing and thylakoids in peanut leaf. It is a critical shade-tolerant mechanism of plants by modulating the development of chloroplast and formating more numbers of thylakoids and grana as well as the grana lamellae [60]. At the later growth stage, chloroplast senescence happened, and the chloroplast shape changed from elliptical to spherical. Then, the accumulation of starch and the gradual disturbance of thylakoid, including distortion of granular arrangement, is accompanied by an increased number and size of translucent plastoglobuli [61]. The coexistence period of soybean was shorter than peanuts in intercrop** systems (Fig. 1F). Especially peanuts suffer from the heavy shade of maize at the later growth stages (reproductive stages) rather than at the early stages (vegetative stages). Then, intercropped soybean obtained a more extended recovery growth than intercropped peanuts after the maize harvest.
The dry matter accumulation of intercropped soybean decreased at the V5 stage but increased at the R4 stage; in contrast, the dry matter accumulation of intercropped peanuts decreased. The variation of dry matter accumulation was consistent with the corresponding leaf traits (Figs. 2 and 7). The yield advantage of intercrop** is affected by the component crop growth and dry matter partition [62, 63]. The previous study documented that shifts of leaf traits, e.g., LAI, SLW, and chlorophyll, increase photosynthesis and change the accumulation and partition of dry matter [64]. The RGR of intercropped soybean varied from -8.5% to 9.4% during the R4-R6 period; in contrast, the RGR of intercropped peanuts ranged from -22.8% to -36.9% (Fig. 9). The competitive use of resources affects the component crops’ growth and dry matter partition [22]. A less competitive component species usually captures few available resources, produces less dry matter due to weakening growth rates, and allocates less dry matter to the grain than the monoculture [3], ultimately affecting the yield advantages of intercrop**. More dry matter was partitioned to seed for soybean; in contrast, the less dry matter was allocated to seed for peanut (Figs. 7 and S3). Firstly, the different responses of soybean and peanut leaf to shading removal lead to differences in dry matter accumulation. Secondly, the coexistence period of soybean was shorter than peanuts, leading to a more extended recovery growth. Thus, intercropped soybean could allocate more photosynthetic products to grain than monoculture soybean. However, more extended shade periods for peanuts, especially the heavy shading during the late reproductive stages, resulted in less dry matter partitioned to the seed. Therefore, intercrop** shapes the characteristics of dry accumulation and partition of legumes (Fig. 9).
The per-plant grain number of intercropped soybean increased (Table 2), while the per-plant grain number and hundred seeds weight of intercropped peanut decreased (Table 2). Compared with 2017, sufficient rainfall benefited maize growth in 2018 (Fig. 7), which may result in heavy shading on legumes. A more extended coexistence period in peanuts than in soybean (Fig. 1) may limit peanuts’ light interception in intercrop** (Figs. 2 and 5). Then, heavy shade with excessive rainfall from July to August 2018 led to fewer seeds and smaller seeds size of intercropped peanuts (Table 2). Although growth suppression happened due to stronger competition from the more dominant crop, its yield loss is also often reduced due to compensatory effects resulting from changes in morphology and functional traits [7, 65, 66]. Compared with monoculture, more erect leaves, greater specific leaf weight, and prolonged growth duration were reported in intercrop** systems [7, 65, 66]. N addition increased soybean grain number, peanut grain number and hundred seeds weight (Table 2). Notably, a more extended recovery growth of soybean than peanut contributed to yield recovery in intercrop** (Table 3). Although legume can meet about 50–60% of the N requirement for growth through biological nitrogen fixation, reasonable N input helps increases grain yield [67]. Finally, intercropped soybean obtained a net effect ranging from 0.44 to 0.50 t ha−1, but a negative value of net effect ranging from -0.48 to 0.61 t ha−1 was obtained for intercropped peanuts (Fig. 7).
Conclusions
In the current study, intercropped maize obtained a yield advantage by strengthening leaf functional traits and dry matter partition. Namely, intercropped maize increases the leaf chloroplasts, grana, and grana lamellae, increasing chlorophyll and SLW to promote dry matter accumulation. Although the shade of maize in the coexistence period has adverse effects on soybean growth, the leaf functional traits, e.g., LAI, SLW, chlorophyll, and chloroplast, are strengthened in the recovered growth stages. Then, the intercropped soybean obtained a yield advantage. In contrast, the leaf functional trait indicates an irreversible penalty of maize’s heavy shade during the coexistence period on peanut growth can not be compensated during the recovered growth period. Finally, the yield disadvantage happened to intercropped peanuts. The land equivalent ratio and the net effect of maize-soybean intercrop** ranges from 1.38 to 1.60 and 2.22 to 3.99 t ha−1, and from 0.89 to 1.13 and 1.24 to 3.26 t ha−1 for maize-peanut intercrop**. Overall, maize-soybean relay intercrop** obtains a win–win yield advantage, and maize-peanut strip intercrop** achieves a trade-off yield advantage.
Availability of data and materials
The original contributions presented in the study are included in this article. Further inquiries can be directed to the corresponding author.
References
Tscharntke T, Clough Y, Wanger TC, Jackson L, Motzke I, Perfecto I, Vandermeer J, Whitbread A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol Cons. 2012;151:53–9.
Vandermeer JH. The ecology of intercrop**. Cambridge University Press; 1989.
Li L, Sun J, Zhang F, Li X, Yang S, Rengel Z. Wheat/maize or wheat/soybean strip intercrop** I. Yield advantage and interspecific interactions on nutrients. Field Crops Res. 2001;71:123–37.
Li X, Wang Z, Bao X, Sun J, Yang S, Wang P, Wang C, Wu J, Liu X, Tian X, et al. Long-term increased grain yield and soil fertility from intercrop**. Nature Sustainabil. 2021;4(11):943–50.
Trenbath B. R: Intercrop** for the management of pests and diseases. Field Crop Res. 1993;34(3–4):381–405.
Brooker RW, Bennett AE, Cong WF, Daniell TJ, George TS, Hallett PD, Hawes C, Iannetta PPM, Jones HG, Karley AJ. Improving intercrop**: a synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015;206(1):107–17.
Zhang W, Gao S, Li Z, Xu H, Yang H, Yang X, Fan H, Su Y, Fornara D, Li L. Shifts from complementarity to selection effects maintain high productivity in maize/legume intercrop** systems. J Appl Ecol. 2021;58(11):2603–13.
Yong T, Chen P, Dong Q, Du Q, Yang F, Wang X, Liu W, Yang W. Optimized nitrogen application methods to improve nitrogen use efficiency and nodule nitrogen fixation in a maize-soybean relay intercrop** system. J Integr Agric. 2018;17(3):664–76.
Li L, Zhang L, Zhang F: Crop mixtures and the mechanisms of overyielding. In: Encyclopedia of Biodiversity. 2013;2:382–95.
Thilakarathna MS, McElroy MS, Chapagain T, Papadopoulos YA, Raizada MN. Belowground nitrogen transfer from legumes to non-legumes under managed herbaceous crop** systems. A review. Agron Sustain Dev. 2016;36:58.
Jensen ES, Carlsson G, Hauggaard-Nielsen H. Intercrop** of grain legumes and cereals improves the use of soil N resources and reduces the requirement for synthetic fertilizer N: A global-scale analysis. Agron Sustain Dev. 2020;40:5.
Hauggaard-Nielsen H, Gooding M, Ambus P, Corre-Hellou G, Crozat Y, Dahlmann C, Dibet A, von Fragstein P, Pristeri A, Monti M, et al. Pea–barley intercrop** for efficient symbiotic N2-fixation, soil N acquisition and use of other nutrients in European organic crop** systems. Field Crop Res. 2009;113(1):64–71.
Zheng B, Zhou Y, Chen P, Zhang X, Du Q, Yang H, Wang X, Yang F, ** promote N uptake through changing the root spatial distribution, legume nodulation capacity, and soil N availability. J Integr Agric. 2022;21(6):1755–71.
Li Y, Yu C, Cheng X, Li C, Sun J, Zhang F, Lambers H, Li L. Intercrop** alleviates the inhibitory effect of N fertilization on nodulation and symbiotic N2 fixation of faba bean. Plant Soil. 2009;323:295–308.
Fan F, Zhang F, Song Y, Sun J, Bao X, Guo T, Li L. Nitrogen fixation of faba bean (Vicia faba L.) interacting with a non-legume in two contrasting intercrop** systems. Plant Soil. 2006;283:275–86.
Zhang R, Meng L, Li Y, Wang X, Ogundeji AO, Li X, Sang P, Mu Y, Wu H, Li S. Yield and nutrient uptake dissected through complementarity and selection effects in the maize/soybean intercrop**. Food Energy Sec. 2020;10: e282.
Carrera CS, Solis SM, Ferrucci MS, Vega CCR, Galati BG, Ergo V, Andrade FH, Lascano RH. Leaf structure and ultrastructure changes induced by heat stress and drought during seed filling in field-grown soybean and their relationship with grain yield. An Acad Bras Ciênc. 2021;93(4): e20191388.
Fan Y, Chen J, Tan T, Li S, Li J, Wang B, Zhang J, Cheng Y, Wu X, Yang W, et al. Soybean (Glycine max L. Merr.) seedlings response to shading: leaf structure, photosynthesis and proteomic analysis. BMC Plant Biol. 2019;19(1):34.
Wu Y, Gong W, Yang F, Wang X, Yong T, Yang W. Responses to shade and subsequent recovery of soya bean in maize-soya bean relay strip intercrop**. Plant Product Sci. 2016;19(2):206–14.
Hunt R. Relative growth rates. In: basic growth analysis: plant growth analysis for beginners. Dordrecht: Springer Netherlands; 1990. p. 25–34.
Zhu J. W vdW, Anten NPR, Vos J, Evers JB: The contribution of phenotypic plasticity to complementary light capture in plant mixtures. New Phytol. 2015;207(4):1213–22.
Zhang D, Sun Z, Feng L, Bai W, Yang N, Zhang Z, Du G, Feng C, Cai Q, Wang Q, Zhang Y. Maize plant density affects yield, growth and source-sink relationship of crops in maize/peanut intercrop**. Field Crops Res. 2020;257(1):107926.
Fan Y, Chen J, Cheng Y, Raza MA, Wu X, Wang Z, Liu Q, Wang R, Wang X, Yong T, et al. Effect of shading and light recovery on the growth, leaf structure, and photosynthetic performance of soybean in a maize-soybean relay-strip intercrop** system. PLoS ONE. 2018;13(5): e0198159.
Echarte L, Maggiora AD, Cerrudo D, Gonzalez VH, Abbateb P, Cerrudo A, Sadras VO, Calvino P. Yield response to plant density of maize and sunflower intercropped with soybean. Field Crops Res. 2011;121:423–9.
Lichtfouse E. Climate change, intercrop**, pest control and beneficial microorganisms, vol. 2. Netherlands: Springer; 2009.
Yin W, Chai Q, Guo Y, Feng F, Zhao C, Yu A, Hu F. Analysis of leaf area index dynamic and grain yield components of intercropped wheat and maize under straw mulch combined with reduced tillage in arid environments. J Agric Sci. 2016;8(4):26–42.
Jumrani K, Bhatia VS. Identification of drought tolerant genotypes using physiological traits in soybean. Physiol Mol Biol Plants. 2019;25(3):697–711.
Ritchie SW, Hanway JJ. How a corn plant develops [Iowa]. Iowa State University Extension; 1982.
Fehr WR, Caviness CE, Burmood DT, Pennington JS. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971;11:929–31.
Buchaillot ML, Soba D, Shu T, Liu J, Aranjuelo I, Araus JL, Runion GB, Prior SA, Kefauver SC, Sanz-Saez A. Estimating peanut and soybean photosynthetic traits using leaf spectral reflectance and advance regression models. Planta. 2022;255(4):93.
Boote KJ. Growth stages of peanut (Arachis hypogaea L.). Peanut Sci. 1982;9:35–40.
Arnon DI. Copper enzyme in isolated chloroplasts polyphenolox-idase in beta vulgaris. Plant Physiol. 1949;24(1):1–15.
Watson DJ. The physiological basis of variation in yield. Adv Agron. 1952;4:101–45.
Loreau M, Hector A. Partitioning selection and complementarity in biodiversity experiments. Nature. 2001;412:72–6.
Van der Werf W, Zhang L, Li C, Chen P, Feng C, Xu Z, Zhang C, Gu C, Bastiaans L, Makowski D, et al. Comparing performance of crop species mixtures and pure stands. Front Agri Sci Engine. 2021;8(3):481–9.
Rao MR, Willey RW. Evaluation of yield stability in intercrop**: studies on sorghum/pigeonpea. Exp Agric. 1980;16:105–16.
R: A language and environment for statistical computing. R Foundation for Statistical Computing [https://www.R-project.org/].
Agricolae: statistical procedures for agricultural research [https://CRAN.R-project.org/package=agricolae].
Siebers MH, Yendrek CR, Drag D, Locke AM, Acosta LR, Leakey ADB, Ainsworth EA, Bernacchi CJ, Ort DR. Heat waves imposed during early pod development in soybean (Glycine max) cause significant yield loss despite a rapid recovery from oxidative stress. Glob Change Biol. 2015;21:3114–25.
Amanullah. Specific leaf area and specific leaf weight in small grain crops wheat, rye, barley, and oats differ at various growth stages and NPK source. J Plant Nutr. 2015;38(11):1694–708.
Murchie EH, Pinto M, Horton P. Agriculture and the new challenges for photosynthesis research. New Phytol. 2009;181(3):532–52.
Kislyuk IM, Bubolo LS, Paleeva TV, Sherstneva OA. Heat-induced increase in the tolerance of the wheat photosynthetic apparatus to combined action of high temperature and visible light: CO2 fixation, photosynthetic pigments, and chloroplast ultrastructure. Russ J Plant Physiol. 2004;51(4):455–63.
Liu Y, Li X, Liu M, Cao B, Tan H, Wang J, Li X. Responses of three different ecotypes of reed (Phragmites communis Trin.) to their natural habitats: leaf surface micro-morphology, anatomy, chloroplast ultrastructure and physio-chemical characteristics. Plant Physiol Biochem. 2012;51:159–67.
Allen JF, Forsberg J. Molecular recognition in thylakoid structure and function. Trends Plant Sci. 2001;6(7):317–26.
Šesták Z. Limitations for finding a linear relationship between chlorophyll content and photosynthetic activity. Biol Plant. 1966;8:336–46.
Mao LZ, Lu HF, Wang Q, Cai MM. Comparative photosynthesis characteristics of Calycanthus Chinensis and Chimonanthus praecox. Photosynthetica. 2007;45(4):601–5.
Raza MA, Cui L, Khan I, Din AMU, Yang W. Compact maize canopy improves radiation use efficiency and grain yield of maize/soybean relay intercrop** system. Environ Sci Pollut Res. 2021;28:41135–48.
Jiao N, Wang J, Ma C, Zhang C, Guo D, Zhang F, Jensen ES. The importance of aboveground and belowground interspecific interactions in determining crop growth and advantages of peanut/maize intercrop**. Crop J. 2021;9(6):1460–9.
Yang F, Liao D, Wu X, Gao R, Fan Y, Raza MA, Wang X, Yong T, Liu W, Liu J, et al. Effect of aboveground and belowground interactions on the intercrop yields in maize-soybean relay intercrop** systems. Field Crop Res. 2017;203:16–23.
Kusnarta IGM, Farida N, Dulur NWD, Wangiyana W. Additive intercrop** with peanut relay-planted between different patterns of rice rows increases yield of red rice in aerobic irrigation system. Emirates J Food Agri. 2021;33(3):202–10.
Sinclair TR. Wit CTd: Photosynthate and nitrogen requirements for seed production by various crops. Science. 1975;189(4202):565–7.
Li L, Tilman D, Lambers H, Zhang F. Plant diversity and overyielding: insights from belowground facilitation of intercrop** in agriculture. New Phytol. 2014;203(1):63–9.
Jo S, Kang Y, Om K, Cha Y, Ri S. Growth, photosynthesis and yield of soybean in ridge-furrow intercrop** system of soybean and flax. Field Crop Res. 2022;275: 108329.
Wu Y, Gong W, Yang W. Shade inhibits leaf size by controlling cell proliferation and enlargement in soybean. Sci Rep. 2017;7(1):9259.
Yang F, Liao D, Fan Y, Gao R, Wu X, Rahman T, Yong T, Liu W, Liu J, Du J, et al. Effect of narrow-row planting patterns on crop competitive and economic advantage in maize-soybean relay strip intercrop** system. Plant Product Sci. 2017;20(1):1–11.
Wittmann C, Aschan G, Pfanz H. Leaf and twig photosynthesis of young beech (Fagus sylvatica) and aspen (Populus tremula) trees grown under different light regime. Basic Appl Ecol. 2001;2:145–54.
Schiefthaler U, Russell AW, Bolhàr-Nordenkampf HR, Critchley C. Photoregulation and photodamage in Schefflera arboricola leaves adapted to different light environments. Funct Plant Biol. 1999;26(5):485–94.
Campbell SJ, Miller CJ. Shoot and abundance characteristics of the seagrass Heterozostera tasmanica in Westernport estuary (south-eastern Australia). Aquat Bot. 2002;73(1):33–46.
Yao Y, Wang S, Kong Y. Characteristics of photosynthesis mechanism in different peach species under low light intensity. Scientia Agricultura Sinica. 2007;40(4):855–63.
Deng Y, Li C, Shao Q, Ye X, She J. Differential responses of double petal and multi petal jasmine to shading: I. Photosynthetic characteristics and chloroplast ultrastructure. Plant Physiol Biochem. 2012;55:93–102.
Zhang M, Zhang C, Yu G, Jiang Y, Strasser RJ, Yuan Z, Yang X, Chen G. Changes in chloroplast ultrastructure, fatty acid components of thylakoid membrane and chlorophyll a fluorescence transient in flag leaves of a super-high-yield hybrid rice and its parents during the reproductive stage. J Plant Physiol. 2010;167:277–85.
Ren Y, Liu J, Wang Z, Zhang S. Planting density and sowing proportions of maize–soybean intercrops affected competitive interactions and water-use efficiencies on the Loess Plateau China. Eur J Agron. 2016;72:70–9.
Poorter H, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytol. 2012;193(1):30–50.
Gao Y, Duan A, Sun J, Li F, Liu Z, Liu H, Liu Z. Crop coefficient and water-use efficiency of winter wheat/spring maize strip intercrop**. Field Crop Res. 2009;111(1–2):65–73.
Liu X, Rahman T, Yang F, Song C, Yong T, Liu J, Zhang C, Yang W. PAR interception and utilization in different maize and soybean intercrop** patterns. PLoS ONE. 2017;12(1): e0169218.
Dong N, Tang M, Zhang W, Bao X, Wang Y, Christie P, Li L. Temporal differentiation of crop growth as one of the drivers of intercrop** yield advantage. Sci Rep. 2018;8(1):3110.
Salvagiotti F, Cassman KG, Specht JE, Walters DT, Weiss A. Nitrogen uptake, fixation and response to fertilizer N in soybeans A review. Field Crop Res. 2008;108(1):1–13.
Acknowledgements
The authors would like to acknowledge their universities for supporting the research, and thanks also go to the two anonymous reviewers for their patient explanations and constructive suggestions.
Funding
The research was supported by the Agriculture Research System of China (soybean) [CARS-04-PS20] and the National Natural Science Foundation of China (3187101212).
Author information
Contributions
ZDF, PC, XNZ, and TWY conceived and designed the experiment. ZDF, PC, and XNZ performed the statistical analysis. ZDF, PC, XNZ, QD, BCZ, HY, and KL were involved in field data collection. ZDF, PC, XNZ, QD, BCZ, HY, KL, PL, YLL, TP, YSW, XCW, FY, WGL, CS, WYY, and TWY contributed to writing the paper. All the authors agreed to submit the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This article does not contain any studies with human or animal subjects. The current experimental research and field study, including the collection of plant material, comply with relevant institutional, national, and international guidelines and legislation and are used for research and development. Maize [(Zea mays L.) **anyu 335] was provided by Dunhuang Seed Industry Pioneer Improved Seed Company Limited. Soybean [(Glycine max L. Merr.) Nandou 25] and peanut [(Arachis hypogaea Linn.) Tianfu 18]were provided by the Agricultural Science Institute of Soybean in Nanchong, Sichuan province.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
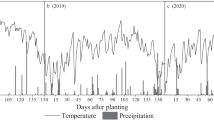
Figure S1. Weather condition of the experimental site. Figure S2. Effects of nitrogen input and crop** system on specific leaf weight (SLW). MM, monoculture maize, MS, monoculture soybean, MP, monoculture peanut, IMS, maize-soybean relay intercrop**, IMP, maize-peanut strip intercrop**. Panels A-B (maize), R1, the silking stage, and R3, the milk stage. Panels C-D (soybean), V5, the fifth trifoliolate stage, and R4, the full pod stage. Panels E-F (peanut), R1, the beginning bloom stage, and R6, the full seed stage. N0, 0 kg N ha-1, N1, 80 kg N ha-1. Data were shown as mean with S.D. Different lower case letter donates significant difference between crop** systems under the same N input (Tukey HSD, p < 0.05). The Tukey HSD values were shown as standard bar above each N treatments. Results of the one-way ANOVA were displayed at the top of each panel, N, N input, C, crop** system, and ‘*’ and ‘ns’ represent significant and insignificant difference at the same growth stage (Tukey HSD, p < 0.05). Figure S3. Effects of nitrogen input and crop** system on aboveground dry matter allocation. MM, monoculture maize, MS, monoculture soybean, MP, monoculture peanut, IMS, maize-soybean relay intercrop**, IMP, maize-peanut strip intercrop**. Panels A-B (maize), V12, the twelfth leaf stage, R1, the silking stage, R3, the milk stage, R6, the maturity stage. Panels C-D (soybean), V5, the fifth trifoliolate stage, R2, the full bloom stage, R4, the full pod stage, R6, the full seed stage, and R8, the maturity stage. Panels E-F (peanut), R1, the beginning bloom stage, R2, the beginning ped stage, R4, the full pod stage, R6, the full seed stage. N0, 0 kg N ha-1, N1, 80 kg N ha-1. Data were shown as mean with S.D.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Fu, Z., Chen, P., Zhang, X. et al. Maize-legume intercrop** achieves yield advantages by improving leaf functions and dry matter partition. BMC Plant Biol 23, 438 (2023). https://doi.org/10.1186/s12870-023-04408-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-023-04408-3