
Abstract
Background
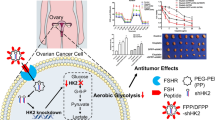
Most ovarian cancers are diagnosed at advanced stages characterized by abdominal dissemination and frequently exhibit chemoresistance. Pyruvate dehydrogenase kinase 2 (PDK2) regulates the switch between glycolysis and oxidative phosphorylation and contributes to tumor progression and chemoresistance. Here, we investigated the effects of PDK2 blockade on metabolic reprogramming and cisplatin sensitivity and evaluated the in vivo antitumor effects of PDK2 shRNA in chemoresistant ovarian cancer using retro-inverso follicle-stimulating hormone peptide-modified nanoparticle as carriers.
Methods
The expression of PDK2 was detected by immunohistochemistry, Western blot and real-time PCR. Cell proliferation and apoptosis were detected using CCK-8 and flow cytometry. Cell migration was detected by Transwell assay. Seahorse Analyzer was used to evaluate metabolic changes. The cisplatin-resistant ovarian cancer cells A2780cp were used to establish the mouse model of peritoneal metastatic ovarian cancer.
Results
A higher expression level of PDK2 was observed in chemoresistant ovarian cancer tissues and cell lines and was associated with shorter progression-free survival. PDK2 knockdown inhibited proliferation and migration and promoted apoptosis of both cisplatin-sensitive and cisplatin-resistant ovarian cancer cells. Cisplatin sensitivity was increased even in cisplatin-resistant ovarian cancer cells. Mechanistically, PDK2 knockdown resulted in an increased oxygen consumption rate and decreased extracellular acidification rate, along with reduced lactate production, increased PDHC activity and increased levels of electron transport chain complexes III and V. The metabolism switched from glycolysis to oxidative phosphorylation. Finally, to specifically and effectively deliver PDK2 shRNA in vivo, we formulated a targeted delivery system containing retro-inverso follicle-stimulating hormone peptide as a targeting moiety and polyethylene glycol–polyethylenimine copolymers as carriers. The nanoparticle complex significantly suppressed tumor growth and peritoneal metastasis of cisplatin-resistant ovarian cancer without obvious toxicities.
Conclusions
Our findings showed the link between metabolic reprogramming and chemoresistance in ovarian cancer and provided an effective targeting strategy for switching metabolic pathways in cancer therapy.
Similar content being viewed by others
Background
Ovarian cancer is the most lethal gynecological cancer with a low early detection rate and high recurrence rate. Approximately 80% of patients have entered stage III or IV at the time of diagnosis, with a 5-year survival rate of less than 30% (Narod 2016; Lheureux et al. 2019). Cytoreductive surgery combined with platinum-based chemotherapy is the primary treatment for advanced ovarian cancer. The R0 resection rate of stage IV epithelial ovarian cancer is only 8.1%, making chemotherapy an independent approach (Winter et al. 2008). However, more than 80% of patients with advanced-stage disease undergo chemoresistance and recurrence and have a poor prognosis (Bowtell et al. 2015).
Aerobic glycolysis, or the well-known “Warburg effect”, is a phenomenon in which cancer cells prefer glycolysis to oxidative phosphorylation (OXPHOS) to produce energy, even under aerobic conditions (Warburg et al. 1927; Lunt and Vander Heiden 2011). But there is some evidence that cancers are not inherently aerobically glycolytic, but anaerobically glycolytic driven by a hypoxia-driven Pasteur effect (Zu and Guppy 2004). And OXPHOS is the main ATP supply pathway under normoxia in some cancer cells such as metastatic triple negative breast cancer cells MDA-MB-231 and MDA-MB-468 (Moreno-Sánchez et al. 2014; Pacheco-Velázquez et al. 2018). The active mitochondrial metabolism is supported by β-oxidation of free fatty acids in hepatoma and cervical cancer cells, which is necessary for ATP demands in the process of cell proliferation (Rodríguez-Enríquez et al. 2015). Nevertheless, accumulated evidence has demonstrated that cancer cells undergo metabolic reprogramming to achieve rapid proliferation and escape the killing effect of chemotherapeutics. Aberrant reprogrammed energy metabolism plays a key role in the tumor microenvironment, initiation, progression and chemoresistance (**ng et al. 2015; Pavlova and Thompson, 2016; Vander Heiden and DeBerardinis 2017; Bergers and Fendt 2021).
Pyruvate dehydrogenase kinases (PDKs), a class of serine/threonine kinases, negatively regulate the activity of the pyruvate dehydrogenase complex (PDHC) through reversible phosphorylation to restrain the transformation of pyruvate to acetyl-coenzyme A (acetyl-CoA). PDKs’ overexpression regulates the metabolic switch between glycolysis and OXPHOS and contributes to the growth, angiogenesis, metastasis, and chemoresistance of cancer, supporting the potential of targeting PDKs in cancer treatment (Wang et al. 2019; Siu et al. 2020; Venturoli et al. 2021; Yao et al. 2021).
Four PDK isoforms have been identified: PDK1, PDK2, PDK3 and PDK4. Among them, PDK2 shows the broadest tissue distribution and the strongest sensitivity to acetyl-CoA and nicotinamide adenine dinucleotide (NADH) (BOWKER-KINLEY et al. 1998; Baker et al. 2000; Harris et al. 2001). PDK2 overexpression is involved in the proliferation and migration of cancer cells and promotes resistance to cisplatin and paclitaxel (Gang et al. 2015; Roh et al. 2016; Sun et al. 2017; He et al. 2018; Hu et al. 2019; Liang et al. 2020). An analysis from TCGA database suggested a positive correlation between higher PDK2 expression and shorter event-free survival or overall survival (Hu et al. 2019; Cui et al. 2020). Inhibition or knockdown of PDK2 could regulate metabolism and reverse chemoresistance of cancer cells using dichloroacetate (DCA) or RNA interference (RNAi) (Roh et al. 2016; Sun et al. 2017; Kitamura et al. 2021). However, the side effects, weak antitumor activity and excessive drug dose limit the clinical application of DCA (Wang et al. 2021).
Additionally, there remain some challenges in terms of stability and transfection efficiency of in vivo delivery for RNAi blocking PDK2. The nanotechnology-based carriers exhibit the potential to overcome these difficulties (Shi et al. 2017; **n et al. 2017; Mohammadinejad et al. 2020). Moreover, the modification of targeted ligands on nanocarriers facilitates the drugs to be delivered into specific lesions (Aghamiri et al. 2019). Peptide-modified delivery system holds great potential because of their high biocompatibility, specific targeting, chemical variety and low toxicity (Tesauro et al. 2019). The binding peptides of follicle-stimulating hormone (FSH) receptor could be appropriate ligands for ovarian cancer (Zhang et al. 2009; Fan et al. 2014; Wang et al. 2018).
In this study, we examined the effects of PDK2 blockade on glucose metabolism and cisplatin sensitivity in ovarian cancer cells. Moreover, to specifically and effectively deliver PDK2 shRNA to ovarian cancer in vivo, we formulated a targeted delivery system containing retro-inverso FSH peptide as a targeting moiety and polyethylene glycol (PEG)-polyethylenimine (PEI) copolymers as carriers. We further investigated the antitumor effects of this targeting strategy for PDK2 in chemoresistant ovarian cancer via the mouse xenograft models of peritoneal metastasis.
Results
Expression of PDK2 in ovarian cancer tissues and cell lines
To evaluate the relevance of PDK2 in ovarian cancer, we first analyzed 185 cases of ovarian cancer from GEO datasets. A higher expression level of PDK2 was observed in ovarian cancer tissues than in normal tissues (Fig. 1A) and was associated with shorter progression-free survival (PFS) (Fig. 1B). Furthermore, our immunohistochemical staining data revealed higher PDK2 protein levels in chemoresistant ovarian cancer tissues (n = 11) than in chemosensitive ovarian cancer tissues (n = 31) (Fig. 1C). Compared with A2780 ovarian cancer cells, the cisplatin-resistant counterpart A2780cp cells showed higher levels of PDK2 protein according to our Western blot data (Fig. 1D). Together, these data suggested a relationship between higher PDK2 expression and chemoresistance in ovarian cancer.

Expression of PDK2 in ovarian cancer tissues and cell lines. A Expression of PDK2 in ovarian cancer and normal tissues of the GEO database (GSE26712). **P < 0.01. B Kaplan–Meier survival curve delineating PFS between the low and high PDK2 expression groups from the GEO database (GSE26712). C Representative images of immunohistochemical staining for PDK2 in ovarian cancer tissues (200X). 1–2, chemosensitive ovarian cancer tissues. 3–4, chemoresistant ovarian cancer tissues. D Expression of PDK2 protein in A2780, A2780cp, Caov3, HEY, SKOV3 and ES2 ovarian cancer cell lines via Western blot
Effects of PDK2 knockdown on the cisplatin sensitivity of ovarian cancer cells
To clarify the effects of PDK2 knockdown on cisplatin sensitivity, we examined cell proliferation and apoptosis in cisplatin-sensitive and cisplatin-resistant ovarian cancer cells. Both A2780 and A2780cp cells were transfected with PDK2 shRNA or scramble shRNA for 24 h before cisplatin treatment. As a result, the expression of PDK2 was strongly silenced at the mRNA and protein levels (Fig. 2A, B). Cell growth was inhibited by PDK2 knockdown in A2780 and A2780cp cells. Notably, the inhibitory effects of cisplatin were significantly enhanced even in cisplatin-resistant cells. The IC50 of cisplatin was substantially reduced after PDK2 knockdown (12.5 μg/ml versus 5 μg/ml) (Fig. 2C, D). Accordingly, the proportions of apoptotic cells increased in the cisplatin-treated A2780 and A2780cp cells at 24 h after transfection with PDK2 shRNA (Fig. 2E, F). The cisplatin sensitivity of ovarian cancer cells was substantially enhanced by PDK2 knockdown.

Effects of PDK2 knockdown on the cisplatin sensitivity of ovarian cancer cells. A PDK2 protein expression in A2780 and A2780cp cells by Western blot. B PDK2 mRNA expression in A2780 and A2780cp cells by real-time PCR. Both A2780 and A2780cp cells were transfected with PDK2 shRNA or scramble shRNA. The viability of A2780 (C) and A2780cp (D) cells measured by CCK-8 assays. Cells transfected with scramble shRNA were used as controls. Both PDK2-depleted and control cells were treated with a concentration gradient of cisplatin for 24 h. The apoptosis of A2780 (E) and A2780cp (F) cells was analyzed using flow cytometry. Both PDK2-depleted and control cells were treated with 5 μg/ml cisplatin for 24 h. ***P < 0.001, **P < 0.01
PDK2 knockdown inhibited the migration of ovarian cancer cells in vitro
Next, to further investigate the effects of PDK2 knockdown on the malignant biobehaviors of cisplatin-resistant ovarian cancer cells, we examined migration via Transwell assays. As shown in Fig. 3, migration was decreased in the PDK2-depleted A2780 and A2780cp cells compared with the control cells transfected with scramble shRNA. Notably, PDK2 knockdown and cisplatin treatment exhibited strong inhibitory effect on migration, even in cisplatin-resistant ovarian cancer cells. In vitro data showed that PDK2 knockdown in ovarian cancer cells suppressed malignant biobehaviors and increased cisplatin sensitivity.

Effects of PDK2 knockdown on the migration of ovarian cancer cells. Both A2780 and A2780cp cells were transfected with PDK2 shRNA or scramble shRNA and then treated with 5 μg/ml cisplatin for 24 h. Representative microscopy images and cell numbers of A2780 (A, C) and A2780cp (B, D) cells. **P < 0.01, *P < 0.05
PDK2 knockdown reprogrammed glucose metabolism in cisplatin-resistant ovarian cancer cells
As important rate-limiting enzymes in glycolysis, PDKs play pivotal roles in regulating glycolysis and OXPHOS. We explored the changes in metabolism after PDK2 knockdown in cisplatin-resistant ovarian cancer cells by Seahorse analysis. After treatment with cisplatin and PDK2 shRNA, the non-glycolytic acid production, glycolysis and glycolytic capacity were decreased (Fig. 4A–D), whereas the ATP production, basal respiration and maximum respiration were increased in cisplatin-resistant A2780cp ovarian cancer cells (Fig. 4E– H). Moreover, the effects of cisplatin on metabolism were significantly enhanced by PDK2 knockdown. We further investigated the effects of PDK knockdown on lactate production. Reduced levels of lactate were found in the supernatants from the PDK2-depleted A2780cp cells compared with the control cells transfected with scramble shRNA (Fig. 4I). Reversely, PDHC activity was increased in the PDK2-depleted A2780cp cells (Fig. 4J). Notably, these metabolic changes were accompanied by increased levels of electron transport chain complexes III and V in the PDK2-depleted A2780cp cells, which was detected via Western blot (Fig. 4K). Taken together, these data suggested that PDK2 knockdown promoted a shift from glycolysis to OXPHOS in cisplatin-resistant ovarian cancer cells as well as restored cisplatin sensitivity.

PDK2 knockdown induced a switch from glycolysis to OXPHOS. A The ECAR measured by Seahorse analysis in cisplatin-resistant A2780cp ovarian cancer cells. Cells were transfected with PDK2 shRNA and then treated with 5 μg/ml cisplatin for 24 h. Quantification of non-glycolytic acid production (B), glycolysis (C) and glycolytic capacity (D). E The OCR measured by Seahorse analysis in A2780cp cells. Quantification of basal respiration (F), ATP production (G) and maximum respiration (H). I Lactate level in supernatants. J PDHC activity in A2780cp cells. A2780cp cells were transfected with PDK2 shRNA (PDK2shRNA group) or scramble shRNA (control group) for 24 h. Then, both PDK2-depleted A2780cp cells and control cells were treated with 5 μg/ml cisplatin for 24 h (cis + PDK2shRNA group vs. cisplatin group). K The levels of electron transport chain complexes in the PDK2-depleted A2780cp cells detected via Western blot. ***P < 0.001, **P < 0.01, and *P < 0.05
PDK2 knockdown suppressed tumor growth and peritoneal metastasis of cisplatin-resistant ovarian cancer
The difficulties of specific and stable delivery in vivo limit the clinical application of RNA interference drugs. We previously developed FSH peptide-modified nanoparticles that can effectively deliver drugs in vivo (Zhang et al. 2009). In this study, we synthesized retro-inverso FSH β 33–53 peptide-conjugated PEG-PEI (DFPP), and then PDK2 shRNA plasmids were loaded into the copolymers. The 1H-NMR spectra of DFPP exhibited the peaks at 3.5–3.6 ppm (from PEG), 2.4–3.0 ppm (from PEI) and 7.0–7.2 ppm (from FSH peptide) (Fig. 5A). The copolymers containing PDK2 shRNA showed spherical shapes under transmission electron microscopy (Fig. 5B). The average DLS particle size of DFPP-PDK2 with an N/P ratio of 25 was 140.6 ± 29.6 nm, and the zeta potential value was − 30.4 ± 1.8 mV.

Characterization of nanoparticle complexes. A 1H-NMR spectroscopy of DFPP copolymer. B Transmission electron microscopy images of PP-PDK2 and DFPP-PDK2. Bar, 100 nm
Then, we used this drug carrier to deliver PDK2 shRNA in vivo and investigated the effects of PDK2 knockdown on tumor growth and metastasis of cisplatin-resistant ovarian cancer. Since ovarian cancer tends to peritoneal disseminate and rapidly accumulate ascetic fluid, a mouse model of peritoneal metastatic ovarian cancer was used in this study. A2780cp cells were injected intraperitoneally into female nude mice to establish the model. Cisplatin, DFPP-PDK2 and the combination of cisplatin and DFPP-PDK2 were administrated, respectively. The changes in abdominal circumference during the study and the number of tumor foci at the end point were measured to reflect the tumor burdens. As shown in Fig. 6, the growth of the abdominal circumference was significantly delayed by the combined administration of DFPP-PDK2 and cisplatin. The number of tumor foci at the end point was the lowest and the inhibition rate was 62.5% in the combination group. Consistent with this finding, the expressions of the proliferation marker Ki67 and the antiapoptotic B-cell lymphoma-2 (Bcl-2) protein obviously decreased in the combination group (Fig. 6E, F). Together, these data showed that DFPP-PDK2 significantly suppressed peritoneal dissemination of chemoresistant ovarian cancer without obvious toxicities.

PDK2 knockdown suppressed tumor growth and peritoneal metastasis of cisplatin-resistant ovarian cancer in vivo. The nude mouse model of cisplatin-resistant peritoneal metastatic ovarian cancer was intraperitoneally injected with cisplatin, DFPP-PDK2 and the combination of cisplatin and DFPP-PDK2 twice a week for 3 weeks. A Representative images of peritoneal tumor foci. B The number of tumor foci at the end point. C The changes of abdominal circumference of the mice. D HE staining in tumor xenografts. Ki67 (E) and Bcl-2 (F) expression via immunohistochemistry in tumor xenografts. (1) control group. (2) cisplatin group. (3) DFPP-PDK2 group. (4) cis + DFPP-PDK2 group. Bars, 50 μm. ***P < 0.001, **P < 0.01, and *P < 0.05
Discussion
Most ovarian cancer patients are diagnosed at an advanced stage and present abdominal dissemination and rapidly accumulating ascetic fluid. Moreover, acquired chemoresistance is a challenge for the majority of these patients.
In contrast to normal cells, cancer cells, fibroblasts and macrophages in the tumor-associated microenvironment prefer aerobic glycolysis (DeBerardinis et al. 2007; **ng et al. 2015). But there are also some cancer cell types that mainly rely on mitochondrial metabolism for ATP supply (Moreno-Sánchez et al. 2014). Cancer cells obtain ATP and intermediates via aerobic glycolysis and/or mitochondrial metabolism to meet the requirements of proliferation and biosynthesis as well as the demands for a beneficial microenvironment (Gatenby and Gillies, 2004; Rodríguez-Enríquez et al. 2015). The untreated cisplatin-resistant ovarian cancer cells A2780cp exhibited a high level of glycolysis in our study. Recent researches have shown that modulating glycolysis could regulate the metastasis and chemosensitivity of ovarian cancer cells (Shi et al. 2018; Tian et al. 2021). PDK2, a primary gatekeeper of glycolysis, contributes to tumor progression and chemoresistance and could be an effective target for cancer treatment (Anwar et al. 2021). Suppression of PDK2 can shift glucose utilization and metabolism and then inhibit tumor growth (Byun et al. 2015). In this study, we investigated the link between metabolic reprogramming and chemoresistance and evaluated the antitumor effects of targeting PDK2 in chemoresistant ovarian cancer.
PDK2 can be blocked by synthetic small-molecule inhibitors and PDHC reaction products. PDK2 inhibition activates PDHC, resulting in a reduction in the levels of end products and further inhibition of PDK2 (Rahman et al. 2016; Shi and McQuibban, 2017). We designed shRNA plasmids to suppress PDK2 expression in vitro and explored metabolic reprogramming after PDK2 knockdown. Cisplatin-resistant ovarian cancer cells showed a lower ECAR and higher OCR after PDK2 knockdown, along with reduced lactate production and increased PDHC activity. The metabolism switched from glycolysis to mitochondrial OXPHOS. As a result, cisplatin sensitivity was restored, cell proliferation and invasion were decreased, and apoptosis was increased. PDK2 blockade could increase the activity of PDHC, regulate the glycolytic phenotype and lactate production, and then induce cell apoptosis or death (Gang et al. 2014, 2015; Hui et al. 2017).
PDK2 is more sensitive to DCA, a small-molecule inhibitor of PDK family members, than the other three isoforms, and this drug has been used in clinical trials for cancer treatment (BOWKER-KINLEY et al. 1998; Tataranni and Piccoli 2019). However, the low efficiency of suppressing tumor growth and systemic toxicity limit its clinical use (Kaufmann et al. 2006; Deuse et al. 2014; Stacpoole et al. 2019; Wang et al. 2021). Other chemical compounds that can inhibit PDK2 activity, such as 2-chloropropionate, also show toxicity (Yount et al. 1982). Because of the ubiquitous tissue distribution of PDK2, precise targeting of PDK2 inhibitors is necessary for their clinical application.
RNAi can efficiently block the genes of interest in vitro and could be a potential tool to target PDK2 expression. In this study, FSH receptor-mediated nanocarrier was used to deliver PDK2 shRNA in vivo. FSH receptor is mainly expressed in ovaries and could be a specific targeting site for ovarian cancer (Perales-Puchalt et al. 2017). The FSH β 33–53 peptide containing natural l-amino acids is able to bind specifically to the FSH receptor and used as targeting moiety in drug delivery system in our previous study (Zhang et al. 2009). However, instability due to protease degradation limited the application of natural peptides as intracellular targets. D-counterparts of L-configuration peptides have inherent proteolytic hyperstability and high binding affinity (Tesauro et al. 2019; Kannan et al. 2020). Retro-inverso FSH peptide containing D-amino acids was used as a targeting moiety to deliver gene drugs into FSH receptor-positive ovarian cancer cells, and showed high affinity and strong stability in our previous and present studies (Zhang et al. 2018). The in vivo data also suggested that PDK2 blockade via this delivery system significantly suppressed peritoneal dissemination of chemoresistant ovarian cancer without obvious toxicities.
This study still has some limitations. Tumors are always heterogeneous, and the expression of PDK2 and FSH receptor varies in different cancer cells. The efficiency of targeted delivery by the FSH receptor-mediated system could be limited. Different histological types of ovarian cancer might have different responses to PDK2 blockade. The effects of PDK2 in different histological types of ovarian cancer need to be further studied.
Conclusions
PDK2 blockade switched glycolysis to OXPHOS in cisplatin-resistant ovarian cancer cells, along with inhibition of cell proliferation and migration as well as increase of cell apoptosis. Retro-inverso FSH peptide-conjugated nanoparticles effectively delivered PDK2 shRNA in vivo and then suppressed peritoneal dissemination of chemoresistant ovarian cancer. This study showed the link between metabolic reprogramming and chemoresistance in ovarian cancer and provided an effective in vivo targeting strategy for switching metabolic pathways in cancer therapy.
Methods
Cell culture and treatment
The A2780 ovarian adenocarcinoma cell line and its cisplatin-resistant counterpart, the A2780cp cell line, were obtained from Professor Yu Yinhua (M.D. Anderson Cancer Center, Houston, TX, USA). The cells were maintained in a 37 °C incubator with 5% CO2. The culture medium was RPMI 1640 containing 10% fetal bovine serum (FBS) (Gibco, South America) and 1% penicillin–streptomycin. For transfection, cells were treated with shRNA plasmid and Lipofectamine 2000 (Invitrogen, USA). Four shRNA sequences of PDK2 were screened and the sequence of 5ʹ-ACATAGAGCACTTCAGCAA-3ʹ was selected due to its high efficiency. The scramble shRNA was used as control. Then, both PDK2-depleted cells and control cells were treated with a concentration gradient of cisplatin (Sigma, USA) for 24 h.
Western blot
Cells were treated with lysis buffer for 30 min and then centrifuged at 20,000 rpm for 15 min at 4 °C. The supernatant was collected, and the protein concentration was detected using a bicinchoninic acid assay (Beyotime, China). Total protein was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membranes. Then, the membranes were incubated with primary antibodies, including PDK2 antibody (Abcam, UK), total OXPHOS human antibody cocktail (Abcam, UK) and β-actin antibody (CST, USA), and HRP-conjugated secondary antibody (Beyotime, China) was added. The protein bands were imaged by chemiluminescence using the ImageQuant LAS4000 system (GE Healthcare Life Sciences, USA).
Real-time PCR
Total RNA was extracted from cells using TRIzol buffer, and then, reverse transcription was performed. Afterward, the reverse transcription products were analyzed by real-time PCR via an ABI PRISM 7900 system (Applied Biosystems, USA). The primers were synthesized by the BioTNT Company (China). The primer sequences were as follows. PDK2: 5ʹ- CGGCTGGCTAACACAATG-3ʹ and 5'-TCCTCAGGGCTCTTATTTTC-3ʹ. GAPDH: 5'GGGAAGGTGAAGGTCGGAGT-3ʹ and 5ʹGGGGTCATTGATGGCAACA-3ʹ. Relative mRNA expression levels were calculated using the 2− ΔΔCT method.
Immunohistochemistry
Human tissues were collected from ovarian cancer patients admitted to the Obstetrics and Gynecology Hospital of Fudan University, which was approved by the Ethics Committee at Obstetrics and Gynecology Hospital of Fudan University. For immunohistochemistry, tissues were fixed in 4% paraformaldehyde, embedded in paraffin and sectioned. The tissue sections were sequentially incubated with primary antibodies, including PDK2 antibody (Abcam, UK), Ki-67 antibody (Abcam, UK) and Bcl-2 antibody (Abcam, UK), and HRP-conjugated secondary antibody. Diaminobenzidine and hematoxylin were used to stain tissues, and then, images were obtained under light microscopy.
Cell viability assay
A CCK-8 kit (Do**do, Japan) was used to assess cell viability. Cells (3 × 103) were seeded in 96-well plates. After treatment with PDK2 shRNA and cisplatin for 24 h, cell viability was examined according to the manufacturer’s instructions.
Cell migration assay
Transwell chambers (Corning, USA) were used to assess cell migration in vitro. After treatment with PDK2 shRNA and cisplatin for 24 h, cells were seeded in the upper Transwell inserts in serum-free medium. The bottom 24-well plate contained medium supplemented with 10% FBS. After culture for 24 h, the cells remaining inside the inserts were removed with cotton swaps. The cells that migrated to the lower surface of the inserts were fixed with 4% paraformaldehyde, stained with crystal violet and counted under light microscopy.
Cell apoptosis assay
Cell apoptosis was examined using a PE-Annexin V apoptosis detection kit (BD Biosciences, USA). After treatment with PDK2 shRNA and cisplatin for 24 h, cells were repeatedly washed with cold PBS and then resuspended in binding buffer at a density of 1 × 106 cells/ml. Next, 5 μl of PE-Annexin V and 5 μl of 7-aminoactinomycin D (7-AAD) were sequentially added to binding buffer for 15 min. Cells were detected and analyzed by FACStation cytometry (BD Biosciences, USA). PE-Annexin V-positive and 7-AAD-negative cells were considered apoptotic cells.
Seahorse metabolic assay
The extracellular acidification rate (ECAR) and oxygen consumption rate (OCR) of treated cells were determined by a Seahorse Bioscience XF96 Analyzer (Agilent, USA) according to the manufacturer’s protocol. Briefly, 1 × 105 cells were seeded in an XF96 culture plate. Glycolysis stress and mitochondrial stress tests were performed. The glycolysis, glycolytic capacity and glycolytic reserve were detected by adding glucose, oligomycin and 2-deoxyglucose (2-DG). The basal values before addition of glucose were non-glycolytic acid production. The basal respiration, proton leak, maximum respiration and spare respiratory capacity were detected by adding oligomycin, carbonylcyanide-p-trifluoromethoxyphenylhydrazone (FCCP), rotenone and antimycin A.
Lactate detection
Cells were cultured in phenol red-free RPMI 1640 after treatment with the indicated drugs for 24 h. Then, the culture medium was collected to measure lactate levels using a lactate assay kit (BioVision, USA) according to the manufacturer’s protocol. The detection value was normalized to the total protein level of cells.
PDHC activity measurement
PDHC activity was measured by a PDH Enzyme Activity Microplate Assay Kit (Abcam, UK). Samples were loaded onto the microplates coating with anti-PDHC antibody and incubated at room temperature. After 3 h, 200 μL assay solution was added into each well. Absorbance at OD450 nm was measured according to the manufacturer’s protocol.
Preparation and characterization of copolymer-containing PDK2 shRNA
Retro-inverso FSH β 33–53 peptide-modified PEG-PEI copolymers were prepared and conjugated with shRNA plasmids as previously described (Zhang et al. 2018). The conjugation of the copolymer was confirmed by 1H nuclear magnetic resonance (1H-NMR) spectroscopy (Bruker BioSpin International, Switzerland). The dynamic light scattering (DLS) particle size, zeta potential and morphology were examined by Malvern Zetasizer Autosizer (Malvern, UK) and transmission electron microscope (JEOL, Ltd., Japan). The retro-inverso FSH peptide-modified PEG-PEI copolymers containing PDK2 shRNA (named as DFPP-PDK2) with an nitrogen to phosphorus (N/P) ratio of 25 were used in next experiments.
Tumor model of peritoneal metastatic ovarian cancer
A total of 1 × 107 A2780cp cells were injected intraperitoneally into the right lower quadrant of 5-week-old female BALB/c nude mice (SLAC Laboratory, Shanghai, China) to establish the model of peritoneal metastatic ovarian cancer. The mice were randomly divided into 4 groups with 6 mice per group using simple random number. All mice were included in this study. Cisplatin, DFPP-PDK2 and the combination of cisplatin and DFPP-PDK2 were injected intraperitoneally twice a week for 3 weeks. The control group was injected with saline. The dose of DFPP-PDK2 was 2.5 mg/kg body weight, and that of cisplatin was 2.5 mg/kg body weight. A first investigator was aware of the treatment group allocation and administered the treatment. A second investigator performed the measurement procedure. The abdominal circumference was measured during the study, and the number of tumor foci was counted at the end point. The animal experiments followed the principles for laboratory animal use and care and with approval from the Institutional Animal Care and Use Committee of SLAC.
Statistical analysis
Student’s t test and one-way analysis of variance were used. P values < 0.05 were considered significant. Statistical Package for the Social Sciences (SPSS) version 16.0 and GraphPad Prism 6.0 were used for data analyses.
Availability of data and materials
Not applicable.
Abbreviations
- OXPHOS:
-
Oxidative phosphorylation
- PDKs:
-
Pyruvate dehydrogenase kinases
- PDHC:
-
Pyruvate dehydrogenase complex
- Acetyl-CoA:
-
Acetyl-coenzyme A
- NADH:
-
Nicotinamide adenine dinucleotide
- DCA:
-
Dichloroacetate
- RNAi:
-
RNA interference
- FSH:
-
Follicle-stimulating hormone
- PEG:
-
Polyethylene glycol
- PEI:
-
Polyethylenimine
- Bcl-2:
-
B-cell lymphoma-2
- FBS:
-
Fetal bovine serum
- SDS-PAGE:
-
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
- ECAR:
-
Extracellular acidification rate
- OCR:
-
Oxygen consumption rate
- 2-DG:
-
2-Deoxyglucose
- FCCP:
-
Carbonylcyanide-p-trifluoromethoxyphenylhydrazone
- 1H-NMR:
-
1H nuclear magnetic resonance
- DLS:
-
Dynamic light scattering
- PFS:
-
Progression-free survival
References
Aghamiri S, Mehrjardi KF, Shabani S, Keshavarz-Fathi M, Kargar S, Rezaei N (2019) Nanoparticle-siRNA: a potential strategy for ovarian cancer therapy? Nanomedicine 14:2083–2100
Anwar S, Shamsi A, Mohammad T, Islam A, Hassan MI (2021) Targeting pyruvate dehydrogenase kinase signaling in the development of effective cancer therapy. Biochim Biophys Acta Rev Cancer 1876:188568
Baker JC, Yan X, Peng T, Kasten S, Roche TE (2000) Marked differences between two isoforms of human pyruvate dehydrogenase kinase. J Biol Chem 275:15773–15781
Bergers G, Fendt S (2021) The metabolism of cancer cells during metastasis. Nat Rev Cancer 21:162–180
Bowker-Kinley MM, Davis IW, Wu P, Harris AR, Popov MK (1998) Evidence for existence of tissue-specific regulation of the mammalian pyruvate dehydrogenase complex. Biochem J 329:191–196
Bowtell DD, Böhm S, Ahmed AA, Aspuria P, Bast RC, Beral V et al (2015) Rethinking ovarian cancer II: reducing mortality from high-grade serous ovarian cancer. Nat Rev Cancer 15:668–679
Byun JK, Choi YK, Kang YN, Jang BK, Kang KJ, Jeon YH et al (2015) Retinoic acid-related orphan receptor alpha reprograms glucose metabolism in glutamine-deficient hepatoma cells. Hepatology 61:953–964
Cui L, Cheng Z, Liu Y, Dai Y, Pang Y, Jiao Y et al (2020) Overexpression of PDK2 and PDK3 reflects poor prognosis in acute myeloid leukemia. Cancer Gene Ther 27:15–21
DeBerardinis RJ, Mancuso A, Daikhin E, Nissim I, Yudkoff M, Wehrli S et al (2007) Beyond aerobic glycolysis: Transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. Proc Natl Acad Sci 104:19345–19350
Deuse T, Hua X, Wang D, Maegdefessel L, Heeren J, Scheja L et al (2014) Dichloroacetate prevents restenosis in preclinical animal models of vessel injury. Nature 509:641–644
Fan L, Chen J, Zhang X, Liu Y, Xu C (2014) Follicle-stimulating hormone polypeptide modified nanoparticle drug delivery system in the treatment of lymphatic metastasis during ovarian carcinoma therapy. Gynecol Oncol 135:125–132
Gang BP, Dilda PJ, Hogg PJ, Blackburn AC (2014) Targeting of two aspects of metabolism in breast cancer treatment. Cancer Biol Ther 15:1533–1541
Gang H, Dhingra R, Lin J, Hai Y, Aviv Y, Margulets V et al (2015) PDK2-mediated alternative splicing switches Bnip3 from cell death to cell survival. J Cell Biol 210:1101–1115
Gatenby RA, Gillies RJ (2004) Why do cancers have high aerobic glycolysis? Nat Rev Cancer 4:891–899
Harris RA, Huang B, Wu P (2001) Control of pyruvate dehydrogenase kinase gene expression. Adv Enzyme Regul 41:269–288
He Z, Li Z, Zhang X, Yin K, Wang W, Xu Z et al (2018) MiR-422a regulates cellular metabolism and malignancy by targeting pyruvate dehydrogenase kinase 2 in gastric cancer. Cell Death Dis 9:505
Hu T, Yu S, Li Y, Ren H, Ning Q, Wang J et al (2019) PDK2 induces cisplatin-resistance in lung adenocarcinoma via transcriptional regulation of CNNM3. J Drug Target 27:460–465
Hui S, Ghergurovich JM, Morscher RJ, Jang C, Teng X, Lu W et al (2017) Glucose feeds the TCA cycle via circulating lactate. Nature 551:115–118
Kannan S, Aronica P, Ng S, Gek LD, Frosi Y, Chee S et al (2020) Macrocyclization of an all-d linear α-helical peptide imparts cellular permeability. Chem Sci 11:5577–5591
Kaufmann P, Engelstad K, Wei Y, Jhung S, Sano MC, Shungu DC et al (2006) Dichloroacetate causes toxic neuropathy in MELAS: A randomized, controlled clinical trial. Neurology 66:324–330
Kitamura S, Yamaguchi K, Murakami R, Furutake Y, Higasa K, Abiko K et al (2021) PDK2 leads to cisplatin resistance through suppression of mitochondrial function in ovarian clear cell carcinoma. Cancer Sci 112:4627–4640
Lheureux S, Braunstein M, Oza AM (2019) Epithelial ovarian cancer Evolution of management in the era of precision medicine. CA A Cancer J Clin 69:280–304
Liang Y, Hou L, Li L, Li L, Zhu L, Wang Y et al (2020) Dichloroacetate restores colorectal cancer chemosensitivity through the p53/miR-149-3p/PDK2-mediated glucose metabolic pathway. Oncogene 39:469–485
Lunt SY, Vander Heiden MG (2011) Aerobic glycolysis: meeting the metabolic requirements of cell proliferation. Annu Rev Cell Dev Bi 27:441–464
Mohammadinejad R, Dehshahri A, Sagar MV, Zahmatkeshan M, Tavakol S, Makvandi P et al (2020) In vivo gene delivery mediated by non-viral vectors for cancer therapy. J Control Release 325:249–275
Moreno-Sánchez R, Marín-Hernández A, Saavedra E, Pardo JP, Ralph SJ, Rodríguez-Enríquez S (2014) Who controls the ATP supply in cancer cells? Biochemistry lessons to understand cancer energy metabolism. Int J Biochem Cell Biol 50:10–23
Narod S (2016) Can advanced-stage ovarian cancer be cured? Nat Rev Clin Oncol 13:255–261
Pacheco-Velázquez SC, Robledo-Cadena DX, Hernández-Reséndiz I, Gallardo-Pérez JC, Moreno-Sánchez R, Rodríguez-Enríquez S (2018) Energy metabolism drugs block triple negative breast metastatic cancer cell phenotype. Mol Pharm 15:2151–2164
Pavlova NN, Thompson CB (2016) The emerging hallmarks of cancer metabolism. Cell Metab 23:27–47
Perales-Puchalt A, Svoronos N, Rutkowski MR, Allegrezza MJ, Tesone AJ, Payne KK et al (2017) Follicle-stimulating hormone receptor is expressed by most ovarian cancer subtypes and is a safe and effective immunotherapeutic target. Clin Cancer Res 23:441–453
Rahman MH, Jha MK, Kim J, Nam Y, Lee MG, Go Y et al (2016) Pyruvate dehydrogenase kinase-mediated glycolytic metabolic shift in the dorsal root ganglion drives painful diabetic neuropathy. J Biol Chem 291:6011–6025
Rodríguez-Enríquez S, Hernández-Esquivel L, Marín-Hernández A, El HM, Gallardo-Pérez JC, Hernández-Reséndiz I et al (2015) Mitochondrial free fatty acid β-oxidation supports oxidative phosphorylation and proliferation in cancer cells. Int J Biochem Cell Biol 65:209–221
Roh J, Park JY, Kim EH, Jang HJ, Kwon M (2016) Activation of mitochondrial oxidation by PDK2 inhibition reverses cisplatin resistance in head and neck cancer. Cancer Lett 371:20–29
Shi G, McQuibban GA (2017) The mitochondrial rhomboid protease parl is regulated by pdk2 to integrate mitochondrial quality control and metabolism. Cell Rep 18:1458–1472
Shi J, Kantoff PW, Wooster R, Farokhzad OC (2017) Cancer nanomedicine: progress, challenges and opportunities. Nat Rev Cancer 17:20–37
Shi J, Ye J, Fei H, Jiang SH, Wu ZY, Chen YP et al (2018) YWHAZ promotes ovarian cancer metastasis by modulating glycolysis. Oncol Rep 41:1101–1112
Siu MKY, Jiang Y, Wang J, Leung THY, Ngu SF, Cheung ANY et al (2020) PDK1 promotes ovarian cancer metastasis by modulating tumor-mesothelial adhesion, invasion, and angiogenesis via α5β1 integrin and JNK/IL-8 signaling. Oncogenesis 9:24
Stacpoole PW, Martyniuk CJ, James MO, Calcutt NA (2019) Dichloroacetate-induced peripheral neuropathy. Int Rev Neurobiol 145:211–238
Sun H, Zhu A, Zhou X, Wang F (2017) Suppression of pyruvate dehydrogenase kinase-2 re-sensitizes paclitaxel-resistant human lung cancer cells to paclitaxel. Oncotarget 8:52642–52650
Tataranni T, Piccoli C (2019) Dichloroacetate (DCA) and cancer: an overview towards clinical applications. Oxid Med Cell Longev 2019:1–14
Tesauro D, Accardo A, Diaferia C, Milano V, Guillon J, Ronga L et al (2019) Peptide-based drug-delivery systems in biotechnological applications: recent advances and perspectives. Molecules. https://doi.org/10.3390/molecules24020351
Tian M, Chen X, Li L, Wu H, Zeng D, Wang X et al (2021) Inhibition of AXL enhances chemosensitivity of human ovarian cancer cells to cisplatin via decreasing glycolysis. Acta Pharmacol Sin 42:1180–1189
Vander Heiden MG, DeBerardinis RJ (2017) Understanding the Intersections between Metabolism and Cancer Biology. Cell 168:657–669
Venturoli C, Piga I, Curtarello M, Verza M, Esposito G, Venuto S et al (2021) Genetic perturbation of pyruvate dehydrogenase kinase 1 modulates growth, angiogenesis and metabolic pathways in ovarian cancer xenografts. Cells-Basel 10:325
Wang P, Fan Y, Lu L, Liu L, Fan L, Zhao M et al (2018) NIR-II nanoprobes in-vivo assembly to improve image-guided surgery for metastatic ovarian cancer. Nat Commun. https://doi.org/10.1038/s41467-018-05113-8
Wang J, Qian Y, Gao M (2019) Overexpression of PDK4 is associated with cell proliferation, drug resistance and poor prognosis in ovarian cancer. Cancer Manag Res 11:251–262
Warburg O, Wind F, Negelein E (1927) The metabolism of tumors in the body. J Gen Physiol 8:519–530
Winter WE, Maxwell GL, Tian C, Sundborg MJ, Rose GS, Rose PG et al (2008) Tumor residual after surgical cytoreduction in prediction of clinical outcome in stage iv epithelial ovarian cancer: a gynecologic oncology group study. J Clin Oncol 26:83–89
**n Y, Huang M, Guo WW, Huang Q, Zhang LZ, Jiang G (2017) Nano-based delivery of RNAi in cancer therapy. Mol Cancer 16:134
**ng Y, Zhao S, Zhou BP, Mi J (2015) Metabolic reprogramming of the tumour microenvironment. FEBS J 282:3892–3898
Yao S, Shang W, Huang L, Xu R, Wu M, Wang F (2021) The oncogenic and prognostic role of PDK1 in the progression and metastasis of ovarian cancer. J Cancer 12:630–643
Yount EA, Felten SY, O’Connor BL, Peterson RG, Powell RS, Yum MN et al (1982) Comparison of the metabolic and toxic effects of 2-chloropropionate and dichloroacetate. J Pharmacol Exp Ther 222:501–508
Zhang X, Chen J, Zheng Y, Gao X, Kang Y, Liu J et al (2009) Follicle-stimulating hormone peptide can facilitate paclitaxel nanoparticles to target ovarian carcinoma in vivo. Cancer Res 69:6506–6514
Zhang M, Zhang M, Wang J, Cai Q, Zhao R, Yu Y et al (2018) Retro-inverso follicle-stimulating hormone peptide-mediated polyethylenimine complexes for targeted ovarian cancer gene therapy. Drug Deliv 25:995–1003
Zu XL, Guppy M (2004) Cancer metabolism: facts, fantasy, and fiction. Biochem Bioph Res Co 313:459–465
Wang X, Shen X, Yan Y, Li H (2021) Pyruvate dehydrogenase kinases (PDKs) an overview toward clinical applications. Biosci Rep. https://doi.org/10.1042/BSR20204402
Acknowledgements
The authors thank the Department of Chemistry, Fudan University, for kindly assisting with nanoparticle characterization.
Funding
This work was supported by the National Natural Sciences Foundation of China (Grant No. 82172747) and the Shanghai Medical Center of Key Programs for Female Reproductive Diseases (Grant No. 2017ZZ01016).
Author information
Authors and Affiliations
Contributions
MZ and MD performed the experiments and wrote the first draft. YMW, XLQ and GLL contributed to the data analysis and explanation. CJX and XYZ designed the study and revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Ethics Committee at Obstetrics and Gynecology Hospital of Fudan University and the Institutional Animal Care and Use Committee of SLAC.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, M., Du, M., Qi, X. et al. Retro-inversion follicle-stimulating hormone peptide-modified nanoparticles for delivery of PDK2 shRNA against chemoresistant ovarian cancer by switching glycolysis to oxidative phosphorylation. Cancer Nano 13, 23 (2022). https://doi.org/10.1186/s12645-022-00129-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12645-022-00129-8