
Abstract
Improving rice quality remains a crucial breeding objective, second only to enhancing yield, yet progress in quality improvement lags behind yield. The high temperature and ripening conditions in Southern China often result in poor rice quality, impacting hybrid rice production and utilization. Therefore, to address this challenge, analyzing the molecular basis of high-quality traits is essential for molecular design breeding of high-quality hybrid rice varieties. In this study, we investigated the molecular basis of grain shape, amylose content, gel consistency, gelatinization temperature, and aroma, which influence rice quality. We discovered that quality related alleles gs3, GW7TFA, gw8, chalk5, Wxb, ALKTT, and fgr can enhance rice quality when applied in breeding programs. Polymerization of gs3, GW7TFA, gw8, and chalk5 genes improves rice appearance quality. The gs3 and GW7TFA allele polymerization increasing the grain’s length-width ratio, adding the aggregation of gw8 allele can further reducing grain width. The chalk5 gene regulates low chalkiness, but low correlation to chalkiness was exhibited with grain widths below 2.0 mm, with minimal differences between Chalk5 and chalk5 alleles. Enhancing rice cooking and eating quality is achieved through Wxb and ALKTT gene polymerization, while introducing the fgr(E7) gene significantly improved rice aroma. Using molecular marker-assisted technology, we aggregated these genes to develop a batch of indica hybrid rice parents with improved rice quality are obtained. Cross-combining these enhanced parents can generate new, high-quality hybrid rice varieties suitable for cultivation in Southern China. Therefore, our findings contribute to a molecular breeding model for grain quality improvement in high-quality indica hybrid rice. This study, along with others, highlights the potential of molecular design breeding for enhancing complex traits, particularly rice grain quality.
Similar content being viewed by others


Introduction
Rice breeding aims to enhance yield and quality, its two crucial objectives. Heterosis utilization has significantly boosted rice yield in recent years. As living standards continue to rise, so does consumer demand for high-quality rice, making it essential for rice breeders to collaboratively improve yield and quality comprehensively (** taste: the molecular discovery of rice genes improving grain size, shape and quality. J Genet Genomics 42:597–599" href="/article/10.1186/s12284-024-00719-7#ref-CR10" id="ref-link-section-d158731556e676">2015).
Rice quality includes milling quality, appearance quality, eating and cooking quality (ECQ) and nutritional quality. Grain shape, chalkiness and transparency are important evaluation indexes of rice appearance quality. Cooking and eating quality are affected by amylose content (AC), gel consistency (GC), and gelatinization temperature (GT). High-quality rice typically exhibits long, thin grains, low chalkiness, medium AC, and strong aroma. Consequently, molecular design breeding efforts have primarily targeted grain shape, AC, GC, GT and aroma content to develop high-quality rice.
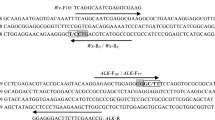
Southern China is the primary region for hybrid rice cultivation, but high temperatures during grain filling stages often compromise the production of high-quality rice (Smith et al. 1997; Preiss and Sivak 1998; Huang et al. 2013; Calingacion et al. 2010; Yang et al. 2023a), necessitating the rejuvenation of parental lines in some early elite hybrid rice combinations to meet current market demands through grain shape improvement. In the early stage, we conducted a genome-wide association study and allelic functional nucleotide polymorphisms analysis of quality trait genes revealed that ALK, FGR1, FLO7, GL7/GW7, GLW7, GS2, GS3, ONAC129, OsGRF8, POW1, WCR1, and Wx were associated with the genetic enhancement of rice quality traits in Southern China. An analysis of 13 crucial rice quality genes, including the fragrance gene fgr, indicated that only a few varieties, such as Gui516, Gui569, Gui721, Ryousi, Rsimiao, Rbasi, and Yuehui9802, possessed multiple superior alleles in Southern China (Yang et al. To accelerate genoty** and molecular marker-assisted (MAS) breeding for high-quality rice, we developed PARMS markers for seven major quality-control genes (GS3, GW7, GW8, Chalk5, Wx, ALK, and FGR), following the method described in the Materials and Methods section. Ten PARMS sets were designed to detect different alleles of these seven genes, with primer sequences and distinguishable SNPs listed in Supplementary Table S1. We validated these markers by genoty** the 214 rice varieties or parental lines from our breeding germplasm collection. Results in Supplementary Figure S1 and Supplementary Table S2 confirmed that all ten PARMS sets specifically differentiated the allelic variations of their target genes. For example, the GW7-90 PARMS set, which detects the A/G SNP of the GW7 gene at the upstream − 90 bp position, clearly distinguished this SNP in 211 out of 214 rice varieties. Similarly, the Chalk5-a PARMS set, detecting the T/C SNP of the Chalk5 gene at the upstream − 485 bp position, unambiguously discerned this SNP in 213 out of 214 rice varieties (Supplementary Table S2, Supplementary Figure S1). These findings demonstrated that the developed PARMS sets reliably and effectively differentiated allelic variations linked to the functional differentiation of these seven quality-control genes. Genetic diversity analysis indicates that these alleles have been stably inherited through long-term natural selection. The high-quality related alleles of gs3, GW7, gw8, chalk5, Wxb, ALKTT, and fgr have gradually been utilized in production. The utilization of grain shape genes is mainly based on gs3 (allele frequency of 0.83), while GW7 (allele frequency of 0.02) and gw8 (allele frequency of 0.08) are less commonly used. Wxb (allele frequency 0.79) and ALKTT (allele frequency 0.72) have also been widely used in high-quality breeding. The application of aroma related fgr (E7) allele (allele frequency of 0.2) in production is gradually increasing (Supplementary Table S3). We genotyped 214 rice varieties or parental lines of hybrid rice for these seven quality control genes and selected 69 elite lines for further genotype-phenotype correlation analyses. These lines represent the backbone parents of indica rice varieties or indica hybrid rice widely cultivated either in the past or at present (Supplementary Table S4). We phenotyped the selected 69 lines for rice grain length and width, AC, ASV, GC, and 2-AP (aroma substance) content (Supplementary Table S4). Upon examining the allelic variation distribution of the seven quality control genes in these 69 elite lines, we observed that the superior gs3 allele, which confers larger and longer rice grains, was widely used in the past in 53/69 (53 out of 69) lines (Supplementary Table S4). This predominance reflects the yield-oriented breeding efforts of the past. The Wxb allele associated with medium-low AC content was also highly represented (45/69), highlighting the continuous focus on improving eating and cooking quality (ECQ) (Supplementary Table S4). The favorable alleles, chalk5 (lower chalkiness) and ALKTT (lower GT) were relatively unbiased in distribution, with 27/69 and 30/69 lines, respectively. However, the favored alleles GW7TFA and gw8Amol, linked to longer and slender rice grains, and fgr, associated with rice aroma, were underrepresented, with only 7/69, 4/69, and 8/69 lines, respectively (Supplementary Table S4). We categorized 69 elite lines into six groups based on the allelic variations of grain size and shape-related genes, GS3, GW7, and GW8 (Fig. 1a-c): I (GS3/GW8/gw7, 15 lines), II (GS3/gw8Amol/gw7, two lines), III (gs3/gw8Amol/gw7, two lines), IV (gs3/GW8/gw7, 43 lines), V (GS3/GW8/GW7TFA, three lines), and VI (gs3/GW8/GW7TFA, four lines). The genotype-phenotype correlation analyses revealed that the GS3 gene primarily controls grain length as lines with the GS3 allele exhibited shorter grains than those with the gs3 allele (I vs. IV and II vs. III, Fig. 1a). Pyramiding the gs3 and GW7TFA alleles further significantly increased grain length (V vs. VI and I vs. VI, Fig. 1a). The gw8Amol allele played a major role in controlling grain width, as all lines carrying this allele displayed slender grains (II vs. I and III vs. IV, Fig. 1b). Additionally, group I (GS3/GW8/gw7) had wider grains than other group (I vs. II, III, IV, VI, Fig. 1b), indicating that GS3 and gw7 genes also contribute minimally to grain width control. Rice lines carrying the gs3 and GW7TFA alleles demonstrated increased grain length, while those with the major gw8Amol allele and the minor GW7TFA allele showed reduced grain width. Consequently, lines possessing either gw8Amol or GW7TFA had a higher grain length-width ratio (3.0-4.3) compared to those with the GS3/GW8/gw7 (group I) genotype (2.0-2.9). Specifically, rice lines with the gs3 and GW7TFA allele combination (group VI) exhibited superior length-width ratios, surpassing 3.5 in the majority of these lines (Fig. 1c, Supplementary Table S4). This combination (group VI) consistently yielded favorable outcomes in grain length, grain width, and length-width ratio (Fig. 1a-c, Supplementary Table S4). Effect analysis of the targeted genes genotype on rice quality. (a), (b) and (c) Effects of the grain type genes gs3, gw8 and GW7 on grain length, grain width and grain length width ratio; (d) and (e) Effect of Chalk5 and grain width on chalkiness. The abscissa is the grain width, A/a > 2.5 mm, B/b = 2.3 ~ 2.49 mm, C/c = 2.0 ~ 2.29 mm, D/d = 1.8 ~ 1.99 mm, E/e < 1.8 mm, Capital letters indicate the high chalkiness allele Chalk5, and lower case letters indicate the low chalkiness allele chalk5; (f), (g), and (h) Effects of Wx and ALK on amylose content (AC), gel consistency (GC) and gelatinization temperature (GT) which was represented by alkali spreading value (ASV), respectively; (i) Effect of fgr on rice fragrance. Significant Z test was annotated in the graph, * P < 0.05, ** P < 0.01. The horizontal lines inside the box represent the median value. The upper side and lower side of the box represent the upper quartile and lower quartile, respectively. The whiskers represent the range of data, and small circles represent outliers The degree and rate of chalkiness in polished rice grains significantly impact both grain appearance quality and ECQ, thus affecting market value. Chalkiness is primarily regulated by the Chalk5 gene, highly correlated with grain width. Our analyses show that when grain width was > 2.5 mm, lines carrying the high chalkiness allele Chalk5 produced grains with substantially higher chalkiness than those with the low chalkiness allele chalk5. For widths between 2.0 and 2.49 mm, the influence of Chalk5 on chalkiness is weak, with smaller widths showing even weaker effects. The chalk5 exhibited low correlation to chalkiness with grain widths below 2.0 mm, with minimal differences between Chalk5 and chalk5 alleles (Fig. 1d-e, Supplementary Table S4). We can also see from the correlation between the average effect of GS3 alleles on grain shape and chalkiness that changes in grain shape greatly affect the formation of chalkiness (Supplementary Table S5). These findings suggest reducing grain width below less than 2.0 mm and is an effective strategy to mitigate chalkiness. Consistent with previous studies (Wang et al. 1995; Cai et al. S2). This further confirms that these alleles are closely related to the quality of rice. Over the past few decades, rice breeders’ efforts have led to the development of elite varieties and parental lines of hybrid rice, significantly contributing to global food security. However, many of these varieties have become obsolete or have been withdrawn from the current market, primarily because their grain quality fails to meet current market preferences. Therefore, updating or improving these elite varieties and parental lines for the current market holds great value for rice breeding. Leveraging our newly acquired knowledge from genotype-phenotype correlation analyses, we aimed to enhance the grain quality of six cytoplastic male sterile (CMS) lines and seven restorer lines using the molecular design breeding strategy described in the Materials and Methods section. By pyramiding the superior alleles, including gs3, GW7TFA, gw8Amol, Wxb, ALKTT, chalk5, and fgr(E7), we anticipate expediting the achievement our breeding objective: to develop high-yielding hybrid rice with premium quality. MeiB, maintainer line of the elite CMS line MeiA with a slender grain phenotype due to carrying the gw8Amol allele, has a grain length-width ratio of ~ 3.08. Introducing the larger and longer grain allele gs3 resulted in the improved line D93, with significantly increased grain length and width. However, the combination of gs3 and gw8Amol led to a slight increase in the grain length-width ratio from 3.08 to 3.17, compared to the original MeiB. Another improved line D99 was developed by pyramiding four favorable alleles: gs3, gw8Amol, GW7TFA and fgr(E7). Compared to D93, D99 exhibited a significant decrease in grain width (from 2.29 mm to 1.70 mm) and a slightly longer grain length became a little longer (from 7.27 mm vs. 7.43 mm), resulting in a significant increase in the grain length-width ratio to 4.37, compared to MeiB’s 3.08 and D93’s 3.17 (Fig. 2a, Supplementary Table S7). Additionally, the fgr(E7) allele in the improved D99 substantially enhanced MeiB’s aroma (Supplementary Table S6). Similarly, the maintainer line YXB of the elite CMS line YXA carrying gs3 and gw8Amol alleles, was improved by adding the GW7TFA allele, which resulted in two new improved lines, D141 and D163, with substantially increased grain length and length-width ratios (Fig. 2b, Supplementary Table S7). Effect of the grain type gene polymerization on appearance quality of rice. (a) Grain shape characteristics of MeiB and its improved lines D93 and D99; (b) Grain shape characteristics of YXB and its improved lines D141 and D163 TeA is an elite CMS line, recognized for its large grain, exceptional plant architecture, and high yield potential, has resulted in its numerous high-yielding hybrid rice combinations being bred and widely cultivated in China. However, TeA’s short and wide grain, high chalkiness grain rate and grain chalkiness degree, and high AC rendered its hybrid rice unsuitable for the current market, TeA to become increasingly obsolete. To address these shortcomings, we employed the TFB donor line, rich in high grain quality genes, to enhance the grain quality of TeB (TeA’s maintainer line). By substituting TeB’s inferior GS3, gw7, and Wxa alleles with TFB’s superior gs3, GW7TFA and Wxb alleles while preserving the superior ALKTT allele of TeB, we developed two improved lines (D123 and D129). The rice quality parameters of both lines were significantly enhanced compared to TeB (Fig. 3a, Supplementary Table S6). Notably, the appearance quality improved, with D123 and D129 exhibiting increases in rice grain length by 35.38% and 30.42%, respectively, and decreases in rice grain width by 30.39% and 30.04%, respectively. Due to this grain length increase and the grain width decrease, the rice grain length-width ratio changed from 2.13 to 4.14 and 3.97 (Fig. 3b), respectively. Additionally, the grain chalkiness degree and chalkiness grain rate significantly reduced, with the grain chalkiness degree decreasing from 45.31 to 1.69% and 2.44% (Fig. 3c), and the chalky grain rate drastically drop** from 100 to 7.90% and 11.58% (Fig. 3d), respectively. Compared to the original TeB, the improved lines D123 and D129 exhibited substantial amelioration in ECQ-related AC and GC. AC decreased from 24.71 to 17.23% and 18.52% (Fig. 3e), while GC increased from 36 mm to 89 mm and 83 mm (Fig. 3f), respectively. Moreover, GT remained at or above 6.5 (Fig. 3g). These results demonstrated that pyramiding the superior gene alleles gs3, GW7TFA, ALKTT and Wxb sufficiently upgraded the grain quality of an obsolete elite parental line. Enhancement of the grain quality in TeB by gene polymerization. (a) The milled grain performance of TeB and its improved lines D123 and D129; (b) Comparison of the rice grain length, grain width and length–width ratio in TeB and its improved lines D123 and D129; (c) and (d) Comparison of the chalkiness characteristics in TeB and its improved lines D123 and D129; (e-g) Comparison of the AC, GC and ASV in TeB and its improved lines D123 and D129, respectively Besides the previously mentioned three improved CMS lines, another three CMS lines (ShanA, II-32A, and BoIIIA) and seven restorer (R) lines (R998, R582, R402, R534, Gui3158, Guiyu9, and Hexin5) were enhanced using the same strategy (Fig. 4, Supplementary Table S7). The data analysis of CMS lines utilizes their corresponding maintainer lines for statistical purposes. These improved parental lines exhibited significant enhancements in grain length, grain width, grain length-width ratio, chalkiness, AC, GC, and GT. The grain length, grain width, and length-width ratio of the improved lines ranged from 7.75 to 9.18 mm (Fig. 4a), 1.73 to 2.09 mm (Fig. 4b), and 4.03 to 4.75 (Fig. 4c), respectively. Their AC fell within the favorable medium range (16.29–18.76%) (Fig. 4d). Moreover, their GC (80–100 mm; Fig. 4e) and GT (ASV grade 6.0–7.0; Fig. 4f) met the standards of high-quality rice. Concurrently, introduction of the fgr(E7) gene also considerably improved the rice’s aroma (Fig. 4i, Supplementary Table S6). Therefore, our results demonstrated that pyramiding gs3, GW7, gw8, ALKTT, chalk5, Wxb, and fgr(E7) alleles represents a broadly applicable strategy for rice quality improvement (Fig. 4j). Enhancement of the grain quality in hybrid rice parents by gene polymerization breeding. (a-c) Comparison of the rice grain length, grain width and length–width ratio in parents and its improved lines (D letter head), respectively; (d-f) Comparison of the AC, GC and GT in parents and its improved lines, respectively; (c) and (h) Comparison of the chalkiness characteristics in parents and its improved lines; (i) Comparison of the 2-AP content in parents and its improved lines; (j) The grain performance of the parents and its improved lines To assess the potential of breeding superior quality hybrid rice using high-quality CMS lines and improved R lines, we performed crosses between them, resulting in nine hybrid rice combinations. Upon analyzing their rice quality, one hybrid rice combination met the premium quality grade 3, and six combinations achieved the premium quality grade 1 of the Chinese standard NY/T 593–2021 (Tang et al. 1999) for cooking rice variety quality (Fig. 5; Table 1). These findings demonstrate that utilizing parental lines enhanced in appearance quality and ECQ significantly increases our likelihood of breeding hybrid rice with superior quality. However, the slender grains of the rice hybrid combination had a negative impact on yield, the increase in grain length-width ratio resulted in a decrease in the thousand grain weight of the rice hybrid combination. The thousand grain weight of these superior quality combinations was generally less than 21 g (Table 1). The pursuit of a high grain length-width ratio would disrupt the balance between yield and quality. Therefore, to achieve both high-quality and high-yield goals in synergy, it is necessary to simultaneously consider multiple traits such as appropriate grain length-width ratio and effective grain number per spike. The combinations of hybrid rice with high quality. (a) The appearance of rice; (b) The characteristics of rice panicles; (c) The plant type characteristics of rice Conventional breeding technology has been instrumental in enhancing rice yield over the past few decades but has certain limitations, such as long breeding cycles, high contingency, and low breeding efficiency. Traditional breeding methods for yield-related traits, like grain size and spike number, rely heavily on visual selection and breeders’ expertise developed over years of experience. easily distinguishable, making successful selection possible. However, traits such as grain ECQ, AC, and resistance to biotic and abiotic stress require laboratory tests or specific conditions for differentiation, making selection more difficult and inefficient, particularly in early segregating populations. Consequently, achieving a balance between yield and grain quality traits is difficult, with high-yielding varieties often exhibiting poor grain quality and good quality varieties having relatively lower grain yield. This issue is particularly prevalent in early hybrid rice, as its CMS parental lines typically have poor grain quality, and genetic improvement of these lines is a lengthy and arduous process. The problem of unbalanced yield and quality somehow restricts the production and promotion of hybrid rice. In contrast, molecular design breeding offers a solution by significantly increasing breeding efficiency and shortening breeding cycles through strategic planning. This approach enables accurate enhancement of current varieties’ shortcomings and rapid aggregation of multiple desirable traits. As more genes controlling crucial agronomic traits are identified and their roles in regulating complex traits are elucidated, precise molecular design breeding in rice is poised to advance rapidly. A recent study by Zeng et al. (Zeng et al. 2001) as the foundation for the penta-primer amplification refractory mutation system (PARMS), a simple and rapid SNP genoty** technology. In this technique, five primers, including a pair of universal fluorescent primers, allele-specific primer pairs, and a shared reverse primers were used to amplify SNP or short indel loci with allele specificity. Subsequently, fluorescence scanning was utilized for genoty** (Zhang et al. 2019; Jun et al. 2020). PARMS markers specific for seven rice quality-related genes (GS3, GW7, GW8, Chalk5, Wx, ALK, and FGR) were developed using allelic variation information of those seven genes gathered from literature. SNPs were identified using BLAST and sequence alignment, and primer sets were designed employing Primer3Plus (https://primer3plus.com). The PARMS marker sequences and corresponding SNP details are listed in Supplementary Table S1. PCR amplification and genotype assays were performed following established protocols (Gao et al. 2021). The developed PARMS markers were then used to genotype 214 indica hybrid rice parental lines and inbred varieties (Supplementary Table S2). Subsequently, 69 were selected for further genotype distribution and genotype-phenotype correlation analyses. These included 16 CMS maintainer lines, 23 CMS restorer lines, and 28 inbred varieties (Supplementary Table S4). For molecular breeding improvement, 13 lines were chosen: six CMS lines TeA, MeiA, YXA, ShanA, II-32 A, BoIIIA and seven restorer lines (R998, R582, R402, R534, Gui3158, Guiyu9, Hexin5) (Supplementary Table S7). All experimental rice plants were cultivated at the Guangxi Academy of Agricultural Sciences farm in Nanning (108°22′ E, 22°48′ N) during regular rice planting seasons. Evaluation of the grain width, grain length, length-width ratio and chalky kernels was performed according to the National Standards of the People’s Republic of China (GB/T17891-1999) (Tang et al. 1999). Mature and dried seeds were shelled, and 100 randomly selected brown rice grains were scanned using a SC-G Automatic Seed Test and 1000-grain Weight Analyzer (Wanshen Testing Technology Co., LTD, Hangzhou, China) to measure the grain length, width, and length-width ratio. The average values were calculated from these 100 grains, with the length-width ratio determined as the grain length divided by its width. Additionally, 100 fully filled polished rice grains were randomly selected to assess grain chalkiness and chalkiness rate using a MICROTEH Scanner (MRS-9600TFU2L) and a Wanshen SC-E Rice Appearance Quality Analyzer (Wanshen Testing Technology Co., LTD, Hangzhou, China). Mature rice grains were harvested, air-dried, and stored at room temperature for three months. For ECQ evaluation, ~ 150 g of grains were de-husked using a huller (SDL-A; CNRRI, Hangzhou, China) and milled using a JMJ-100 rice miller (CNRRI, Hangzhou, China). The ECQ parameters, including AC, GC, GT (evaluated as alkaline spreading value, ASV) were measured according to the methods reported previously (Huang et al. 2013). The grain ECQ parameter was analyzed independently in two consecutive years with three technical repeats for each test. The headspace solid phase microextraction (HS-SPME) method was employed to extract the total volatile components of rice (Kataoka et al. 2000). Refined rice, retrieved from a -80 °C refrigerator, was swiftly ground into refined rice powder. Then 3 g of the refined rice powder was transferred into a 15 ml headspace bottle, followed by complete sealing and insertion of an extraction head into the headspace bottle. Extraction was conducted in a water bath at 60 ℃ for 45 min with the headspace bottle containing the extraction head. Subsequently, gas chromatography-mass spectrometry (GC-MS) (Agilent 7890B-5977 A) was used to analyze the contents of 2-AP and other volatile components at 220 °C using the external standard method. Six elite CMS maintainer lines (B lines) and seven elite restorer lines (R lines) of three-line hybrid rice were selected to advance quality improvement. Those lines were used as the recurrent parents, while the elite CMS maintainer line TFB (carrying the favored gs3, GW7TFA, ALKTT and Wxb alleles) and the elite R line GX204 (carrying the favored gs3, GW7TFA, ALKTT, Wxb and fgr(E7) alleles) were chosen as the donor parents for the B lines and R lines, respectively. Crossing, genoty**, selection, and backcrossing were performed from 2015 to 2021 as shown in Supplementary Figure S3. Using molecular marker assisted selection technology combined with backcrossing to introduce high-quality related alleles, each generation selects high-quality strains, and the higher generation self-crosses with homozygous genetic background to breed stable genetic single plants and form high-quality strains, such as MeiB and YXB (Supplementary Figure S4). Subsequently, the improved B lines with stable inheritance of agronomic traits were converted to corresponding CMS lines (A lines). The resulting high-quality A lines were then tested with high-quality R lines including the improved R lines, to breed premium quality hybrid rice. Box-plot combinations is depicted by the genotypic and phenotypic data of 69 elite lines (Supplementary Table S4) in Microsoft’s Excel software. Significance testing be treatment by the Z test module and compare between every group. Basic statistics of genetic diversity including total number of alleles, and polymorphism information content (PIC) at each allele locus according to the formula PIC = 1-∑pi2 (Nei 1973). Display of phylogenetic tree using ggtree package in R language.Result
Development of PARMS Markers for the Major Quality-Control Genes, GS3, GW7, GW8, Chalk5, Wx, ALK and FGR
Genotype-Phenotype Correlation Analyses among the 69 Selected Elite Parental Lines or Varieties

Improve the Grain Quality of Elite Parental Lines of Hybrid Rice by Molecular Design Breeding



Improved Elite Parental Lines Significantly Increase the Possibility to Breed Hybrid Rice with Superior Quality

Discussion
Plant Materials
Measurements of the Grain Quality
Determination of the Volatile Compound 2-acetyl-1-pyrroline (2-AP) Content
Rational Design Breeding Procedure
Data Analysis
Data Availability
All datasets generated for this study are included in the article/Supplementary file.
References
Bai X, Luo L, Yan W, Kovi MR, Zhan W, ** a pleiotropic quantitative trait locus qGL7. BMC Genetics, 11:16
Bradbury LM, Fitzgerald TL, Henry RJ, ** Q, Waters DL (2005) The gene for fragrance in rice. Plant Biotechnol J 3:363–370
Cai XL, Wang ZY, **ng YY, Zhang JL, Hong MM (1998) Aberrant splicing of intron 1 leads to the heterogeneous 5’ utr and decreased expression of waxy gene in rice cultivars of intermediate amylose content. Plant J 14:459–465
Calingacion M, Laborte A, Nelson A, Resurreccion A, Concepcion JC, Daygon VD, Mumm R, Reinke R, Dipti S, Bassinello PZ, Manful J, Sophany S, Lara KC, Bao J, **e L, Loaiza K, El-Hissewy A, Gayin J, Sharma N, Rajeswari S, Manonmani S, Rani NS, Kota S, Indrasari SD, Habibi F, Hosseini M, Tavasoli F, Suzuki K, Umemoto T, Boualaphanh C, Lee HH, Hung YP, Ramli A, Aung PP, Ahmad R, Wattoo JI, Bandonill E, Romero M, Brites CM, Hafeel R, Lur HS, Cheaupun K, Jongdee S, Blanco P, Bryant R, Thi LN, Hall RD, Fitzgerald M (2014) Diversity of global rice markets and the science required for consumer-targeted rice breeding. PLoS ONE 9:e85106
Dai Z, Lu Q, Luan X, Ouyang L, Guo J, Liang J, Zhu H, Wang W, Wang S, Zeng R, Liu Z, Zhang Z, Zhu X, Zhang G (2016) Development of a platform for breeding by design of CMS restorer lines based on an SSSL library in rice (oryza sativa l). Breed Sci 66:768–775
Fan C, **ng Y, Mao H, Lu T, Han B, Xu C, Li X, Zhang Q (2006) GS3, a major qtl for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet 112:1164–1171
Gao Z, Zeng D, Cheng F, Tian Z, Guo L, Su Y, Yan M, Jiang H, Dong G, Huang Y, Han B, Li J, Qian Q (2011) ALK, the key gene for gelatinization temperature, is a modifier gene for gel consistency in rice. Journal of Integrative Plant Biology, 53:756–765
Gao J, Liang H, Huang J, Qing D, Wu H, Zhou W, Chen W, Pan Y, Dai G, Gao L, Deng G (2021) Development of the PARMS marker of the TAC1 gene and its utilization in rice plant architecture breeding. Euphytica, 217
Gong J, Miao J, Zhao Y, Zhao Q, Feng Q, Zhan Q, Cheng B, **a J, Huang X, Yang S, Han B (2017) Dissecting the genetic basis of grain shape and chalkiness traits in hybrid rice using multiple collaborative populations. Mol Plant 10:1353–1356
Harberd NP (2015) Sha** taste: the molecular discovery of rice genes improving grain size, shape and quality. J Genet Genomics 42:597–599
Hu J, Wang Y, Fang Y, Zeng L, Xu J, Yu H, Shi Z, Pan J, Zhang D, Kang S, Zhu L, Dong G, Guo L, Zeng D, Zhang G, **e L, **ong G, Li J, Qian Q (2015) A rare allele of GS2 enhances grain size and grain yield in rice. Mol Plant 8:1455–1465
Huang R, Jiang L, Zheng J, Wang T, Wang H, Huang Y, Hong Z (2013) Genetic bases of rice grain shape: so many genes, so little known. Trends Plant Sci 18:218–226
Jeon JS, Ryoo N, Hahn TR, Walia H, Nakamura Y (2010) Starch biosynthesis in cereal endosperm. Plant Physiol Biochem 48:383–392
Jun L, **g H, Yidan O, Hui L, Jianhua Z, Chao M, Meng H, Lei W, **ghua X, Yanyu Y, Xu L (2020) A direct PCR-based SNP marker-assisted selection system (D-MAS) for different crops. Mol Breeding 40:9
Kataoka H, Lord HL, Pawliszyn J (2000) Applications of solid-phase microextraction in food analysis. J Chromatogr A 880:35–62
Kovach MJ, Calingacion MN, Fitzgerald MA, Mccouch SR (2009) The origin and evolution of fragrance in rice (Oryza sativa L). Proc Natl Acad Sci USA 106:14444–14449
Li Y, Fan C, **ng Y, Jiang Y, Luo L, Sun L, Shao D, Xu C, Li X, **ao J, He Y, Zhang Q (2011) Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet 43:1266–1269
Li Y, Fan C, **ng Y, Yun P, Luo L, Yan B, Peng B, **e W, Wang G, Li X, **ao J, Xu C, He Y (2014) Chalk5 encodes a vacuolar H(+)-translocating pyrophosphatase influencing grain chalkiness in rice. Nat Genet 46:398–404
Liu J, Chen J, Zheng X, Wu F, Lin Q, Heng Y, Tian P, Cheng Z, Yu X, Zhou K, Zhang X, Guo X, Wang J, Wang H, Wan J (2017) GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat Plants 3:17043
Mao H, Sun S, Yao J, Wang C, Yu S, Xu C, Li X, Zhang Q (2010) Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc Natl Acad Sci USA 107:19579–19584
Miura K, Matsuoka M (2015) Rice genetics: control of grain length and quality. Nat Plants 1:15112
Nei M (1973) Analysis of gene diversity in subdivided populations. Proc Natl Acad Sci U S A 70:3321–3323
Preiss J, Sivak MN (1998) Biochemistry, molecular biology and regulation of starch synthesis. Genetic Eng (N Y) 20:177–223
Sano Y (1984) Differential regulation of waxy gene expression in rice endosperm. Theor Appl Genet 68:467–473
Si L, Chen J, Huang X, Gong H, Luo J, Hou Q, Zhou T, Lu T, Zhu J, Shangguan Y, Chen E, Gong C, Zhao Q, **g Y, Zhao Y, Li Y, Cui L, Fan D, Lu Y, Weng Q, Wang Y, Zhan Q, Liu K, Wei X, An K, An G, Han B (2016) OsSPL13 controls grain size in cultivated rice. Nat Genet 48:447–456
Singh N, Kaur L, Sandhu KS, Kaur J, Nishinari K (2006) Relationships between physicochemical, morphological, thermal, rheological properties of rice starches. Food Hydrocoll 20:532–542
Smith AM, Denyer K, Martin C (1997) The synthesis of the starch granule. Annu Rev Plant Physiol Plant Mol Biol 48:67–87
Song XJ, Huang W, Shi M, Zhu MZ, Lin HX (2007) A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet 39:623–630
Tang R et al (1999) GB/T17891-1999 in National Standard of people’s Republic of China. Standards Press of China
Tian Z, Qian Q, Liu Q, Yan M, Liu X, Yan C, Liu G, Gao Z, Tang S, Zeng D, Wang Y, Yu J, Gu M, Li J (2009) Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc Natl Acad Sci USA 106:21760–21765
Wang ZY, Zheng FQ, Shen GZ, Gao JP, Snustad DP, Li MG, Zhang JL, Hong MM (1995) The amylose content in rice endosperm is related to the post-transcriptional regulation of the waxy gene. Plant J 7:613–622
Wang S, Wu K, Yuan Q, Liu X, Liu Z, Lin X, Zeng R, Zhu H, Dong G, Qian Q, Zhang G, Fu X (2012) Control of grain size, shape and quality by OsSPL16 in rice. Nat Genet 44:950–954
Wang S, Li S, Liu Q, Wu K, Zhang J, Wang S, Wang Y, Chen X, Zhang Y, Gao C, Wang F, Huang H, Fu X (2015) The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat Genet 47:949–954
**ng Y, Zhang Q (2010) Genetic and molecular bases of rice yield. Annu Rev Plant Biol 61:421–442
Yang T, Gu H, Yang W, Liu B, Liang S, Zhao J (2023a) Artificially selected grain shape gene combinations in Guangdong simiao varieties of rice (Oryza sativa L). Rice (N Y) 16:3
Yang X, Pan Y, **a X, Qing D, Chen W, Nong B, Zhang Z, Zhou W, Li J, Li D, Dai G, Deng G (2023b) Molecular basis of genetic improvement for key rice quality traits in southern China. Genomics 115:110745
Ye S, Dhillon S, Ke X, Collins AR, Day IN (2001) An efficient procedure for genoty** single nucleotide polymorphisms. Nucleic Acids Res, 29
Ying JZ, Ma M, Bai C, Huang XH, Liu JL, Fan YY, Song XJ (2018) TGW3, a major QTL that negatively modulates grain length and weight in rice. Mol Plant 11:750–753
Zeng D, Tian Z, Rao Y, Dong G, Yang Y, Huang L, Leng Y, Xu J, Sun C, Zhang G, Hu J, Zhu L, Gao Z, Hu X, Guo L, **ong G, Wang Y, Li J, Qian Q (2017) Rational design of high-yield and superior-quality rice. Nat Plants 3:17031
Zhang X, Wang J, Huang J, Lan H, Wang C, Yin C, Wu Y, Tang H, Qian Q, Li J, Zhang H (2012) Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice. Proc Natl Acad Sci USA 109:21534–21539
Zhang W, Tan L, Sun H, Zhao X, Liu F, Cai H, Fu Y, Sun X, Gu P, Zhu Z, Sun C (2019) Natural variations at TIG1 encoding a TCP transcription factor contribute to plant architecture domestication in rice. Mol Plant 12:1075–1089
Zhao DS, Li QF, Zhang CQ, Zhang C, Yang QQ, Pan LX, Ren XY, Lu J, Gu MH, Liu QQ (2018) GS9 acts as a transcriptional activator to regulate rice grain shape and appearance quality. Nat Communication 9:1240
Acknowledgements
We thank Dr Shuangyong Yan from Institute of Crops Sciences, Tian** Academy of Agricultural Sciences for his selfless assistance in the statistical analysis of some experimental data in the present study.
Funding
This research study was supported by grants from the National Natural Science Foundation of China (U20A2032), the Guangxi Key R&D Program (2023AB17113), the National Natural Science Foundation of China (32360495), Special Fund for Basic Scientific Research of Guangxi Academy of Agricultural Sciences (GuiNongKe2020YM06), The Development Fund of Guangxi Academy of Agricultural Sciences (GuiNongKe2021JM100), Guangxi Innovation Driven Development Science and Technology Project (Major science and technology projects)(GuikeAA22068087-4), Independent Innovation Project Guangxi Key Laboratory of Rice Genetics and Breeding, Guangxi Major Science and Technology Innovation Base (2022-36-Z01-ZZ07).
Author information
Authors and Affiliations
Contributions
JG, LG and GFD conceived the original plan for this work and designed the research. JG, LG and WC performed most of the experiments and collected datasets. JH, DQ, YP, HW and WZ performed parts of experiments. JG performed the analyses and wrote the manuscript. GXD, CM, JL and XY reviewed the manuscript and provided inputs to improve the manuscript. GFD and GXD supervised the project and overviewed all aspects of the work. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gao, J., Gao, L., Chen, W. et al. Genetic Effects of Grain Quality Enhancement in Indica Hybrid Rice: Insights for Molecular Design Breeding. Rice 17, 39 (2024). https://doi.org/10.1186/s12284-024-00719-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12284-024-00719-7