

Abstract
Investigative exploration and foraging leading to food consumption have vital importance, but are not well-understood. Since GABAergic inputs to the lateral and ventrolateral periaqueductal gray (l/vlPAG) control such behaviors, we dissected the role of vgat-expressing GABAergic l/vlPAG cells in exploration, foraging and hunting. Here, we show that in mice vgat l/vlPAG cells encode approach to food and consumption of both live prey and non-prey foods. The activity of these cells is necessary and sufficient for inducing food-seeking leading to subsequent consumption. Activation of vgat l/vlPAG cells produces exploratory foraging and compulsive eating without altering defensive behaviors. Moreover, l/vlPAG vgat cells are bidirectionally interconnected to several feeding, exploration and investigation nodes, including the zona incerta. Remarkably, the vgat l/vlPAG projection to the zona incerta bidirectionally controls approach towards food leading to consumption. These data indicate the PAG is not only a final downstream target of top-down exploration and foraging-related inputs, but that it also influences these behaviors through a bottom-up pathway.
Similar content being viewed by others

Introduction
Feeding is one of the most important aspects of an animal’s life. Searching for food, animals must conduct careful exploration to gain access to caloric resources and to become familiar with their environment. Such exploratory activities are composed of a broad range of behaviors that include approach to novel stimuli, pursuit of prey and investigation of the environmental layout1,2,3,4. These processes are thus a critical feature of hunting for prey and also for finding caloric resources.
The neural basis for this vital behavior is not well-understood, but such investigative and exploratory curiosity can be reduced by inactivation of zona incerta (ZI) neurons5,6. Furthermore, activation of GABAergic ZI cells causes both exploratory approach and curiosity towards novelty, predation and eating of non-prey foods (i.e., high fat food)7, indicating that the circuits controlling exploration and food consumption8 may be linked. Another circuit that controls similar exploratory investigation and hunting is the medial preoptic area, as activation of camk2a cells from this region causes pursuit and following of both crickets and even of inedible objects9. Both GABAergic ZI cells and preoptic area camk2a cells project to the midbrain periaqueductal gray (PAG) and activation of both of these projections induces cricket hunting in mice7,9. Activation of the camk2a preoptic projection to the PAG also elicits following of objects and crickets, suggesting there are local PAG cells that can trigger pursuit of prey, foraging and environmental investigative exploration9. Moreover, increases in cricket hunting were observed following activation of PAG inputs originating in the lateral hypothalamus7,9, the bed nucleus of the stria terminalis10, the central amygdala nucleus11 and the basal forebrain12. These data suggested that the PAG is a common downstream target of exploratory foraging and hunting circuits, and suggest the existence of PAG cells that induce food-seeking. Indeed, to this end it has been shown that pharmacological inactivation of the PAG decreases food approach and consumption13.
Activation of diverse upstream PAG inputs from higher brain regions induce hunting7,9,11,12,14, but it is unknown if bottom-up feeding circuits originating in the PAG play a role in this behavior. The PAG is also known to affect higher brain regions in the context of defensive behaviors with bottom-up (midbrain to forebrain) projections15,16. For example, the PAG provides a teaching signal to the amygdala to provide CS-US associations during fear conditioning17. Additionally, amygdala lesions completely blocked fleeing responses evoked by dorsal PAG stimulation18 suggesting that the basolateral amygdala is downstream of the dorsal PAG in mediating such behaviors. These data indicate that the PAG may also influence hunting and foraging by affecting higher brain regions, though such a mechanism has not been described. It is reasonable that the elaborate display of PAG-evoked behavioral responses would be organized by forebrain circuits15,19. In addition, the lateral PAG is also well-positioned to influence reward-seeking through its projections to forebrain sites such as the lateral hypothalamic area20 or the subthalamic ZI19,21. Considering this view, and taking into account that GABAergic PAG activity has been linked to hunting, we hypothesized that these cells control not only hunting, but general exploratory foraging behavior, pursuit of prey and objects as well as approach towards non-prey food sources through a bottom-up projection.
Here, we show that the activity of vgat-expressing GABAergic l/vlPAG cells encode approach towards and consumption of prey and non-prey food. The activity of these cells also bidirectionally controls appetitive approach drive for both food types in mice. Furthermore, activation of l/vlPAG vgat cells induces a wide range of appetitive-related behaviors, such as exploratory foraging and compulsive eating without altering innate defensive behaviors. These data, along with the higher activity displayed by l/vlPAG vgat cells during approach to food indicate an important role for this population in foraging-related approach towards food. We also show that l/vlPAG vgat cells receive inputs from numerous known feeding and exploration nodes, and they project to regions that control eating as well, such as the ZI. Intriguingly, activity in the l/vlPAG vgat projection to the ZI is necessary and sufficient to induce hunting and consumption of non-prey food. Taken together, our data indicate that common pathways control exploratory foraging and pursuit of prey and non-prey foods and that midbrain circuits can influence this vital behavior by acting on higher brain regions.
Results
l/vlPAG vgat cells represent approach and consumption of food
To investigate how l/vlPAG vgat cells encode exploratory approach towards food sources and eating, we injected the vector AAV-DIO-GCaMP6s in the l/vlPAG of vgat-Cre mice to express the genetically encoded calcium indicator GCaMP6s in l/vlPAG vgat cells. This approach allowed us to obtain calcium transients from these cells through implanted miniaturized microscopes (Fig. 1a) during exposure to live crickets and walnuts (Fig. 1b). Eating onset aligned calcium activity shows that cells are often more active prior to eating both food sources (Fig. 1c). To verify if l/vlPAG vgat cells encoded relevant behaviors in this assay, we performed principal component analysis and determined that commonly occurring behaviors, such as approach to food, eating and rearing, appear predominantly in discrete clusters in principal component space (Fig. 1d, e). The clustering silhouette score, measuring quality, is higher than zero for all mice in both assays (Fig. 1d, e), was higher than expected by chance for each individual mouse (Supplementary Fig. 1) and did not correlate with speed (Supplementary Fig. 2), suggesting the specific encoding of feeding-related processes in vgat l/vlPAG cells. Supporting this idea, it was possible to use five-fold cross validated multinomial logistic regression to decode approach to food and eating behaviors from l/vlPAG vgat cell activity above chance levels (Fig. 1f). We next explored if a conserved pattern of neural activation was used to encode these behaviors across cricket and walnut assays. To do so, we co-registered cells recorded in both behavioral assays (Fig. 1g and Supplementary Fig. 3). We then plotted the data in n-dimensional space, where n is the number of neurons, and quantified the degree of clustering of a given behavior across both assays. A scheme illustrating this procedure is shown in Fig. 1h. The left graph shows the eating time points plotted for n = 3 neurons. Dark blue and light blue points correspond to data from the two assays (walnut and cricket). Note that the dark and light blue time points occupy a similar space, and are thus highly overlap**. Consequently, the average distance between walnut eating and cricket eating points is low. Conversely, in the right graph in Fig. 1h the approach points from walnut and cricket assay are more well-separated from each other, and thus the average distance between points from the two assays is higher. Thus, in this analysis, a higher distance between points from the two assays indicates that a particular behavior has a more independent and less conserved representation across the two assays. The actual experimentally-observed data is shown in Fig. 1i. Note that the distance between food approach points is higher than that of eating points. We also analyzed data from both assays in principal component space (Fig. 1j), and show that the average distance between approach cluster points across walnut and cricket assays is greater than that of eating cluster points. Finally, the mean Euclidean distance between n-dimensional behavioral samples and their cluster center across coregistered cells from the cricket and walnut assays is smaller for the eating cluster than the approach cluster, indicating a greater conservation of neural representation for eating than approaching food (Fig. 1k). Taken together, these data show that, both in n-dimensional neuronal space as well as in principal component space, the neural encoding of eating behavior (consummatory) has a more well-conserved representation across assays than the representation of the more variable approach behavior (appetitive) to food.

a Surgery for recording l/vlPAG vgat calcium transients (top). Example l/vlPAG vgat traces (bottom). b Order of assays. c L/vlPAG vgat activity centered at eating onset (cricket assay n = 223 neurons; walnut assay n = 204 neurons). d Representation of behaviors in principal component space (PC) during cricket hunting (see example on left). Clustering quality was measured by silhouette score, which was higher than the chance level of zero (dotted red line) (n = 4 mice; one-sample two-tailed t-test, t-statistic = 2.79, p = 0.059). e Same as d, but for walnut (n = 4 mice; one-one-sample two-tailed t-test, t-statistic = 14.37, p = 0.001). f Behaviors were decoded above chance (red dotted line) (n = 4 mice; one-sample two-tailed t-test, cricket t-statistics: approach = 3.87, p = 0.031; eat = 5.59, p = 0.011; walnut t-statistics: approach = 4.75, p = 0.018, eat = 7.53, p = 0.005). g Cells co-registered across assays. h Mahalanobis distance between points from two clusters that display higher overlap (left panel with light and dark blue points) and two clusters that are well-separated (right panel with light and dark green points). i Mahalanobis distance between approach and eating clusters across the cricket and walnut assays in n-dimensions (n = # of co-registered neurons). The distance between eating clusters across assays is smaller than the distance between approach clusters, indicating the representation of eating is more conserved than food approach (approach sample n = 527 time points, eat sample n = 1958 time points; two-tailed Wilcoxon rank-sum test, z-score = 22.88, p < 0.001). j Same as d, but for co-registered cells, showing conserved representation of behaviors across assays (n = 4 mice; one-one-sample two-tailed t-test, t-statistic = 4.22, p = 0.024). k The distance between individual points and cluster center is smaller in the eating cluster than the approach cluster, indicating the representation of eating is more conserved across assays (approach sample n = 1049 time points, eat sample n = 5492 time points; two-tailed Wilcoxon rank-sum two-tailed test, z-score = 10.35, p < 0.001). ***p < 0.001, **p < 0.01, *p < 0.05, † p = 0.059. Data are presented as mean values +/- SEM. Source data are provided as a Source Data File.
l/vlPAG vgat cells show conserved encoding of consumption across prey and non-prey foods
We next investigated the temporal dynamics of l/vlPAG vgat calcium traces during the cricket and walnut assays. Mice were exposed to the walnut assay once and to the cricket assay twice (cricket 1 and 2) (Fig. 2a). Example activity traces show that these cells are more active prior to eating than during eating (indicated by shaded blue boxes in Fig. 2b). Averaged group data show that neural activity was consistently higher prior to eating onset in both assays (Fig. 2c, d). Accordingly, there were more negatively modulated than positively modulated cells during eating (‘eat -‘ and ‘eat +‘ cells, respectively, in Fig. 2e, f). We then evaluated if vgat cells show consistent eating-related modulation across assays (see example co-registration across days for cricket 1 and cricket 2 assays in Fig. 2g). We used a generalized linear model (GLM) to calculate the eating weight for each individual cell. The example cell in Fig. 2h shows a negative weight of similar magnitude across assays, indicating that l/vlPAG vgat cells may show conserved eating-related modulation. Indeed, eating GLM weights for the entire population of recorded co-registered cells were significantly correlated both across cricket 1 and cricket 2 exposures as well as across cricket and walnut assays (Fig. 2i). Approach and eating GLM weights were not correlated (Supplementary Fig. 2), indicating that ensembles encode these behaviors independently. To further characterize this result, we measured the decrease in df/F during eating for each cell across assays (calculated as post-eating df/F - pre-eating df/F). The example cell in Fig. 2j shows a similar decrease in activity following eating onset. This effect was also observed across the population of recorded cells. Cells classified as negatively modulated eat- cells in the walnut experiment showed a decrease in activity during eating in both walnut and cricket 1 assays. Similarly, cricket 1 eat- cells also showed less activation during consumption of food in the cricket 2 assay (Fig. 2k). We also observed that cricket 1 eat- cells showed a similar pattern of neural activation in cricket 2 during eating (Fig. 2l). Indeed, the activity profiles of co-registered eat- and of eat+ cells in cricket 1 and cricket 2 were highly positively correlated (Fig. 2l). Performing the same analysis across walnut and cricket assays revealed that eat- cells also showed highly correlated activity profiles while eating these two different types of food. Similar results were also seen after performing the same analysis using walnut and cricket 2 assays (Supplementary Fig. 4). Interestingly, eat+ activity profiles were not correlated across walnut and cricket assays, showing that the negative modulation during the consummatory phase is well-conserved across different assays, while positive modulation during eating is specific to the consummatory phase of each food type (Fig. 2m).

a Order of assays. b L/vlPAG vgat cells are more active prior to eating (blue rectangle). c Left: Representative cell showing activation prior to eating cricket. Right: pre-post eating activity of all cells (n = 223 cells; two-tailed Wilcoxon signed rank test, z-value = 4.36, p < 0.001). d Same as c, but for walnut (n = 204 cells; two-tailed Wilcoxon sign rank test, z-value = 3.78, p < 0.001). e Left: More cells were negatively modulated by eating cricket (n = 4 mice; two-one-sample two-tailed t-test, t-statistic=3.15, p = 0.026). Right: Mean traces (± 1 s.e.m.) of all cells modulated by eating. (eat- n = 86 cells, eat+ n = 37 cells). f Same as e, but for walnut (left: n = 4; two-one-sample two-tailed t-test, t-statistic=5.88, p = 0.009; right: eat - n = 110 cells, eat + n = 36 cells). g Cells co-registered in cricket 1 and cricket 2. h General linear model (GLM) weights for representative cell in crickets 1 and 2. i Correlation of eating GLM weight across assays (coregistered cricket n = 78 cells, coregistered walnut n = 74 cells; Spearman correlation, left: p < 0.001, right: p = 0.005). j Example cell co-registered in cricket 1 and 2 shows decreased activity after eating onset in both assays. k (left pair of bars) Eat- walnut cells show decreased activity during eating of both walnut (blue) and cricket 1 (grey) (n = 53 cells; two-tailed Wilcoxon signed rank test, walnut z-value = 2.18, p = 0.029; coregistered cricket z-value = 2.72, p = 0.007) (right pair of bars). Same as the two bars on the left, but for cells co-registered in cricket 1 and 2 (n = 35 cells; two-tailed Wilcoxon signed rank test, cricket 1 z-value = 4.49, p < 0.001; coregistered cricket 2 z-value = 3.44, p = 0.001). l Left: Activity of eat- cells in cricket 1 (light blue). Activity for the same cells co-registered in cricket 2 (grey). Right: Correlation of activity centered at eating onset for eat- (light blue) and eat+ (dark blue) cells co-registered in cricket 1 and 2 (eat- n = 35 cells, eat+ n = 17 cells; Spearman correlation). m Same as l, but for cells co-registered in walnut and cricket 1 (eat - n = 53 cells, eat + n = 15 cells; Spearman correlation; for r-value comparison, Fisher r-to-z transformation, p = 0.004). ***p < 0.001, **p < 0.01, *p < 0.05. All error bars and error bands correspond to +/- SEM. Source data are provided as a Source Data File.
l/vlPAG vgat cell activity is necessary and sufficient for food foraging and consumption
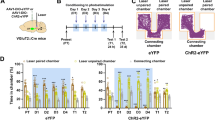
In order to investigate if l/vlPAG vgat cell activity is necessary for appetitive drive and food consumption, we optogenetically inhibited these cells during cricket and walnut exposure. To do so, we expressed the inhibitory opsin Arch in these cells (Fig. 3a, b). Optogenetic inhibition of l/vlPAG vgat cells reduced walnut consumption and successful predation, and also increased the latency to attack the cricket (Fig. 3c, d). We also showed that optogenetic excitation of ChR2-expressing l/vlPAG vgat cells produces the opposite effect. This manipulation increased both hunting and food consumption, decreased the latency to eat in both assays (Fig. 3e, f) and was not influenced by sex differences (Supplementary Fig. 5). Activity of l/vlPAG vgat cells is higher during exploration and approach to food, and is lower during consumption (Fig. 2c–f). We next sought to mimic this pattern of activation optogenetically. To do so, we restricted the optogenetic activation of ChR2-expressing l/vlPAG vgat cells only to exploration and approach epochs, and we turned the laser OFF at consumption onset (Supplementary Fig. 6). This manipulation also increased consumption, and thus shows that optogenetic PAG vgat activation during eating is not necessary to increase consumption (Supplementary Fig. 6). Importantly, neither activation or inhibition of l/vlPAG vgat cells altered locomotion speed in an empty open field (Supplementary Fig. 7c). The observed optogenetic effects were not due to viral spread outside the l/vlPAG or into the dorsal raphe (Supplementary Fig. 8, 9). It is noteworthy that a prior report showed that activation of PAG gad2 cells decreases feeding10. Since both gad2 and vgat are commonly used GABAergic markers, these data appear to be in disagreement. We now show that only 42% of the gad2-expressing PAG cells also co-express vgat (Supplementary Fig. 10). Thus, gad2 and vgat are substantially non-overlap** populations within the PAG, explaining the discrepancy between the current study and the prior report on consumption-suppressing gad2 PAG cells10.

a Left: Scheme showing experimental approach for expressing inhibitory (Arch) and excitatory (ChR2) opsins in l/vlPAG vgat cells. Right: Representative Image showing fiber placement and viral expression in l/vlPAG vgat cells. Similar images were obtained in all 13 animals used. b Experimental timeline. c Optogenetic inhibition of l/vlPAG vgat cells decreases amount of walnut eaten (YFP n = 6, Arch n = 7; right: Wilcoxon rank-sum, z-value = 2.50, p = 0.006; Wilcoxon signed rank (Arch), z-value = −2.20, p = 0.028). d Inhibition of these cells also increases latency to first cricket attack, latency to successful predation and the probability of successful predation (YFP n = 7, Arch n = 8, left: Wilcoxon rank-sum, z-value = −2.03, p = 0.042; Wilcoxon signed rank (Arch), z-value = 2.38, p = 0.013; middle: Wilcoxon rank-sum, z-value = −2.72, p = 0.007; Wilcoxon signed rank (Arch), z-value = 2.52, p = 0.006; right: Wilcoxon rank-sum, z-value = 2.10, p = 0.036; Wilcoxon signed rank (Arch), z-value = −2.25, p = 0.024). e Optogenetic excitation of l/vlPAG vgat cells decreased latency to eat walnut and increased amount walnut eaten (YFP n = 5, ChR2 n = 15, left: Wilcoxon rank-sum, z-value = 1.96, p = 0.049; Wilcoxon signed rank (ChR2), z-value = −3.41, p < 0.001; right: Wilcoxon rank-sum, z-value = −3.14, p = 0.002; Wilcoxon signed rank (ChR2), z-value = 3.41, p < 0.001). f Excitation of ChR2-expressing l/vlPAG vgat cells decreased latency for the first cricket attack and the latency to successful predation. This manipulation also increased the probability of successful predation. (YFP n = 6, ChR2 n = 9, left: Wilcoxon rank-sum, z-value = 3.13, p < 0.001; Wilcoxon signed rank (ChR2), z-value = −2.67, p = 0.008; middle: Wilcoxon rank-sum, z-value = 3.15, p < 0.001; Wilcoxon signed rank (ChR2), z-value = −2.67, p = 0.008; right: Wilcoxon rank-sum, z-value = −3.21, p < 0.001; Wilcoxon signed rank (ChR2), z-value = 2.69, p = 0.007). ***p < 0.001, **p < 0.01, *p < 0.05. Data are presented as mean +/- SEM. Source data are provided as a Source Data File.
Prior studies have shown that activation of other hunting circuits, such as the GABAergic projection from lateral hypothalamus to l/vlPAG, induces both hunting and aggression towards conspecifics14. We thus tested if activation of l/vlPAG vgat cells induces aggression towards other mice, but no behavioral changes were found (Supplementary Fig. 11), indicating that the biting and attack elicited by l/vlPAG vgat activation is specific to predation. These data show that l/vlPAG vgat cell activity is necessary and sufficient for triggering appetitive drive, leading to food approach and consumption of prey and non-prey food types.
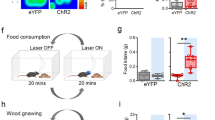
To further characterize the role of l/vlPAG vgat cells in foraging and exploration, we optogenetically activated these cells in a variety of situations. First, we show that activation of ChR2-expressing (Fig. 4a) l/vlPAG vgat cells in an empty chamber induced more rearing, which is a common exploratory behavior (Fig. 4b). To verify if this manipulation would induce more complex exploration we added a climbing ladder to the environment and observed that optogenetic activation decreased the latency to climb (Fig. 4c). To further investigate the variability of context-specific exploratory behaviors, we then exposed the mouse to the holeboard task, which contained holes in the floor that the mouse could explore by performing head dips. Activation of vgat cells consistently increased the number of hole visits in this assay (Fig. 4d). To determine if activity in these cells produced positive motivation, we performed the real time place preference task. Mice displayed a preference for the chamber paired with optogenetic stimulation, indicating that this manipulation may be rewarding or engage positive motivational processes (Fig. 4e). We next reasoned that vgat activation may induce chasing, as hunting is a motivated action. We attached a ball to a long stick and moved the ball in a trajectory that spelled the letters “BG” as previously described9 (Fig. 4f, left panel in upper row; also see Supplementary Movie 1). During blue light delivery, control YFP-expressing mice did not follow the ball (note in Fig. 4f, middle panel in upper row, that the behavioral mouse track, shown in black does not overlap with the ‘BG’ track made by the ball). Strikingly, ChR2-expressing mice closely followed the ball during the light ON period (note in Fig. 4f, right panel in the upper row, that mouse and ball tracks are highly overlap**). Several measurements averaged across mice indicate that vgat stimulation induced following of the ball, as this manipulation decreased the average distance from the ball, increased correlation between mouse and ball position and decreased the angular offset between mouse head direction and ball (Fig. 4f, lower row). These data show that optogenetic stimulation induces context-specific exploration (Fig. 4b–d) as well as approach towards a moving object (Fig. 4f). Considering these results, we next tested if this manipulation would cause directed exploration and climbing to approach a food source that is only accessible by climbing a wire mesh ladder. Indeed, stimulation of vgat cells decreased the latency to climb and bite a walnut and increased time spent near the walnut (Fig. 4g). Vigorous climbing was visible fairly quickly after onset of light stimulation (see Supplementary Movie 2), similar to the rapid induction of cricket hunting following activation of l/vlPAG vgat cells (Supplementary Movie 3). No changes in locomotion were observed in this experimental condition (Supplementary Fig. 12). Following this assay, we tested if vgat activity caused preferential approach to food compared to an object. To do so, we hung a walnut and an object in opposite corners of the same chambers. Both items were outside of the reach of the mouse. Activation of l/vlPAG vgat cells increased the time spent near the walnut corner (see heatmap in Fig. 4h). Additionally, this manipulation also increased the time spent near the walnut, but not the object (Fig. 4h). These optogenetic activation data show that l/vlPAG vgat stimulation produced exploratory foraging (Fig. 4g, h) and positive motivation or preference (as shown in the real time preference task in Fig. 4e). We thus investigated if vgat activation would increase food consumption, even if this behavior is associated with negative consequences, which is a cardinal feature of compulsive eating. We created an environment in which part of the floor was covered by an active shocking grid. To gain access to the walnut, mice would have to endure the negative consequence of exposure to foot shocks (see scheme in Fig. 4i). In both YFP and ChR2-expressing mice, delivery of shocks decreased the amount of walnut consumed. Furthermore, activation of ChR2-expressing vgat l/vlPAG cells rescued the decrease in eating in the presence of the foot shocks. It is worth pointing out that none of these effects were due to analgesia, as excitation of l/vlPAG vgat cells did not alter pain reactions (Supplementary Fig. 13). These data show that vgat stimulation is sufficient to induce eating, even in the presence of aversive consequences—which is a cardinal feature of compulsive eating. We also show that stimulating l/vlPAG vgat cells for a longer period (30 min) induces consumption of a large amount of food (Supplementary Fig. 14), which may also contribute to compulsive feeding symptoms. Taken together, these optogenetic activation data show that l/vlPAG vgat activity can induce exploration, foraging, place preference, chasing of moving objects, proximity to food and certain aspects of compulsive eating.

a Experimental timeline (bottom). b L/vlPAG vgat stimulation increases rearing in an empty box (YFP n = 7, ChR2 n = 7; Wilcoxon rank-sum, z-value = 3.07, p < 0.001) and also c climbing in an environment with a ladder (YFP n = 7, ChR2 n = 7; Wilcoxon rank-sum, z-value = −3.28, p < 0.001). d L/vlPAG vgat activation increased headdips into holes in the holeboard test, and also increased distance traveled. Lower rows: Representative tracks (YFP n = 6, ChR2 n = 6; left: Wilcoxon rank-sum, z-value = −2.81, p = 0.005; right: Wilcoxon rank-sum, z-value = −2.428, p = 0.015). e Left: Behavioral heat maps showing l/vlPAG vgat stimulation induced real-time place preference (YFP n = 6, ChR2 n = 10; Wilcoxon rank-sum, z-value = −3.48, p < 0.001; Wilcoxon signed rank (ChR2), z-value = 3.09, p = 0.002). f A ball was moved on a trajectory spelling “BG” (upper left). L/vlPAG vgat activation decreased distance to ball, increased correlation of the position of mouse and ball and decreased the head angle offset between mouse and ball (YFP n = 7, ChR2 n = 5; Wilcoxon rank-sum; left: z-value = 2.76, p = 0.006; middle: z-value = −2.43, p = 0.015; right: z-value = 2.76, p = 0.006). g L/vlPAG stimulation decreased latency to climb a ladder and then bite the walnut, and increased time spent in the walnut zone (YFP n = 6, ChR2 n = 5; bottom left: Wilcoxon rank-sum, z-value = 2.71, p = 0.007; Wilcoxon signed rank (ChR2), z-value = −2.02, p = 0.043; bottom right: Wilcoxon rank-sum, z-value = −2.65, p = 0.008; Wilcoxon signed rank (ChR2), z-value = 2.02, p = 0.043). h An object and a walnut were suspended in opposite corners (upper left). L/vlPAG vgat activation increased rearing and time spent in the walnut corner, but not the object corner (YFP n = 6, ChR2 n = 5; bottom left: Wilcoxon rank-sum (walnut), z-value = −2.65, p = 0.008; bottom right: Wilcoxon rank-sum (walnut), z-value = −2.65, p = 0.008). i To measure compulsive eating, walnuts were placed on a shocking grid to evaluate if mice endured foot-shocks to access walnuts. L/vlPAG vgat stimulation (shock+laser epoch) increased consumption. (YFP n = 5, ChR2 n = 7; Wilcoxon signed rank (baseline vs. shock) YFP: z-value = 1.75, p = 0.08; ChR2: z-value = 2.37, p = 0.018; Wilcoxon rank-sum (shock+laser), z-value = 2.76, p = 0.01) ***p < 0.001, **p < 0.01, *p < 0.05, † p = 0.08. Data are presented as mean +/- SEM. Source data is in the Source Data File.
Though this work’s focus is on the PAG’s role in appetitive drive and eating behaviors, the l/vlPAG is also known to be a central node for the coordination of innate defensive behaviors22,23,24. Furthermore, it is plausible that food consumption may be inhibited in high anxiety states. These views raise the possibility that higher l/vlPAG vgat activity increases consumption by inhibiting anxiety and other defensive states. We then tested if l/vlPAG vgat activity is sufficient and necessary for defensive states elicited by the anxiogenic elevated plus maze or by a live predatory rat. Bidirectional manipulation of these cells failed to induce changes in open-arm exploration or in the number of risk-evaluation head dips in the elevated plus maze (Supplementary Fig. 15a–d). Similarly, optogenetic excitation or inhibition of l/vlPAG vgat activity also did not alter a wide number of behavioral metrics in the predatory rat exposure assay, including escape speed, percent time freezing and distance to the predator rat (Supplementary Fig. 15e–j). These data indicate that l/vlPAG vgat activity does not modulate defensive states caused by open spaces or predators. To test if l/vlPAG vgat activity is modulated by proximity to innate threats we expressed the calcium indicator GCaMP6s in l/vlPAG vgat cells and obtained recordings of calcium transients during exposure to the elevated plus maze and a live predatory rat (Supplementary Fig. 15k, l). Heatmaps of neural activity show that vgat cells do not consistently encode distance to threat (Supplementary Fig. 15m, n), as higher activity was not displayed consistently either near or far from either threat category. Indeed, l/vlPAG vgat activity was not correlated with distance to either the open arms or the rat (Supplementary Fig. 15o). Taken together, these data suggest it is unlikely that l/vlPAG vgat cells affected eating as an indirect consequence of anxiety suppression.
l/vlPAG vgat cells are bidirectionally connected with feeding circuits
Our data show that l/vlPAG vgat cells regulate food consumption. We thus hypothesized that these cells would be interconnected with other regions known to regulate eating. To test this idea, we injected AAV5-hSyn-FLEX-TVA-P2A-eGFP-2A-oG in the l/vlPAG of vgat-Cre mice (Fig. 5a). Three weeks later, we injected EnvA-dG-rabies-mCherry in the same location (Fig. 5b). Numerous double-labeled starter cells were observed in the l/vlPAG (Fig. 5c). Rabies-infected mcherry-expressing inputs to l/vlPAG vgat cells were seen in several regions known to control hunting, including the medial preoptic area, the central amygdala, the lateral hypothalamic area, the bed nucleus of the stria terminalis and the ZI (Fig. 5d). Interestingly, activation of projections from all these regions to the l/vlPAG induce hunting7,9,11,12,14. These data show that l/vlPAG vgat cells are a common downstream target of diverse circuits that control hunting related behaviors. However, the potential downstream targets of the l/vlPAG vgat cells that may mediate its role in eating have not been identified. We thus next studied if l/vlPAG vgat cells projected to regions that control hunting or food consumption. We injected AAV5-DIO-EFf1a-YFP in the l/vlPAG of vgat-Cre mice (Fig. 6a) and mapped the resulting projection patterns. Projections from l/vlPAG vgat cells were seen in regions known to control feeding, such as the bed nucleus of the stria terminalis and the hypothalamus (Fig. 6c, d). Interestingly, the strongest projection was seen in the ZI. We now identify a previously uncharacterized monosynaptic GABAergic projection from l/vlPAG vgat cells to the ZI (Fig. 6e–g). Though the functional role of this l/vlPAG to ZI projection is unknown, prior data have shown that activation of either ZI vgat cell bodies or of the ZI projection to the l/vlPAG causes hunting, and ZI vgat cells were also activated during food deprivation7. Thus, both anatomical and functional data suggest that the l/vlPAG to ZI projection may be involved in foraging.

a In order to identify inputs to PAG vgat cells, vgat-Cre mice were injected with AAV5-hsyn-FLEX-TVA-P2A-eGFP-2A-oG in the PAG. Three weeks later, mice were injected with EnvA-dG-rabies-mCherry in the same location. b Timeline of viral injections. c Expression of viral vectors in the vlPAG and the lPAG in vgat-Cre mice. Double-labelled GFP (green) and mCherry (red)-expressing vgat PAG cells (shown in yellow in ‘Merge’ panel) show starter cells for monosynaptic retrograde tracing. d mCherry-expressing (red) cells that project to PAG vgat cells in the bed nucleus of the stria terminalis (BST), medial preoptic area (MPA), central amygdala (CeA), Zona incerta anterior and posterior regions (ZIa and ZIp, respectively), lateral hypothalamic area (LHa) and cuneiform nucleus (Cun). e Quantification of retrogradely-labelled cells across brain regions. Structures are ordered from anterior to posterior locations. Note that the Zona incerta (ZI) provides the most prominent input to PAG vgat cells (n = 6807 starter cells and 16,704 retrogradely labeled cells). Anterior-posterior coordinates in mm from Bregma (B) is displayed in the lower left corner of each image. acp anterior commissure, posterior part, aca anterior commissure, anterior part, AH anterior hypothalamus, BST bed nucleus of the stria terminalis, BLA basolateral amygdala, CeA central amygdala, Cun cuneiform nucleus, cp cerebral peduncle, DMH dorsomedial hypothalamus, DR dorsal raphe, HDB nucleus of the horizontal limb of the diagonal band, IC inferior colliculus, LHa lateral hypothalamus area, MPA medial preoptic area, dPAG lPAG and vlPAG dorsal, lateral and ventrolateral periaqueductal gray, respectively, PH posterior hypothalamus, PMd dorsal premammillary nucleus, SNl, SNc, SNr lateral, compact and reticular parts of the substantia nigra, respectively, VMH ventromedial hypothalamus, ZIa and ZIp zona incerta anterior and posterior regions, respectively. Data are presented as mean values +/- SEM. Source data are provided as a Source Data File.

a In order to characterize the anterograde connectivity of l/vlPAG vgat cells, vgat-Cre mice were injected with AAV5-EF1a-DIO-YFP in the PAG. b Expression of YFP in the l/vlPAG of vgat-Cre mice. c YFP axon terminals that project to the bed nucleus of the stria terminalis (BST), d hypothalamus and e zona incerta (ZI). b–e Similar images were obtained in all 5 animals used. f Scheme showing injections used to characterize monosynaptic GABAergic input from PAG vgat cells to the zona incerta. g (Left panel) Representative traces of ex vivo slice recordings of zona incerta neurons following optogenetic excitation of ChR2-expressing axon terminals of vgat l/vlPAG cells. Traces show recordings in the absence of drugs (control), 0.5 µM tetrodotoxin (TTX), 0.5 µM TTX + 100 µM 4-Aminopyridine (4AP) and 25 µM bicuculline (BIC) (from the top to the bottom). g (Right panel) Quantitative analysis of the amplitude of light-evoked inhibitory postsynaptic currents from the zona incerta for the control (n = 12 cells from 5 mice), 0.5 µM TTX (n = 9 cells from 5 mice), 0.5 µM TTX + 100 µM 4-Aminopyridine (n = 9 cells from 5 mice) and 25 µM bicuculline conditions (n = 6 cells from 3 mice). One-way ANOVA test following by a means comparison Tukey test, NS Not Significant, aca anterior commissure, AH anterior hypothalamus, LHa lateral hypothalamus area, VMH ventromedial hypothalamus. Data are presented as mean values +/- SEM. Source data are provided as a Source Data File.
Activity in the ascending projection from l/vlPAG vgat cells to zona incerta can bidirectionally control food consumption
To dissect the activation patterns of the l/vlPAG vgat projection to the ZI, we obtained recordings of calcium transients in this projection. We expressed the calcium indicator GCaMP6s in l/vlPAG vgat cells and implanted fiberoptic cannulae in axons terminating in the ZI (Fig. 7a, b). Fiber photometry recordings show that both in the walnut and the cricket assay, this projection was more active prior to eating onset, as illustrated by representative traces and averaged data (Fig. 7d–g). We also studied if this activation prior to eating could be observed without touching the food. We thus built a U-shaped maze; at the end of each arm, we placed either an object or a walnut behind wire mesh screens (Fig. 7h). Our data show that the l/vlPAG vgat projection to ZI was activated during approach to a walnut, but not an object, even if the walnut was out of reach and thus could not be consumed or touched (Fig. 7i–k). These data show that the l/vlPAG vgat projection to the ZI is activated during approach to food, prior to eating onset, indicating that this bottom-up, midbrain to subthalamic region circuit may have a role in consumption and approach to food. In support of this view, l/vlPAG vgat cells encode proximity to prey (Supplementary Fig. 16f, g).

a (left) Scheme for fiber photometry recording. (right) vgat-Cre mice were injected with a vector encoding cre-dependent GCaMP6s in the l/vlPAG. A fiberoptic cannula was implanted above the zona incerta (ZI) to record calcium transients in l/vlPAG vgat axon terminals in the zona incerta. b Image showing GCaMP6s-expressing l/vlPAG vgat axon terminals in the ZI. Similar images were obtained in all 8 animals used. c Experimental timeline. d Example recording showing that activity in the l/vlPAG vgat projection to zona incerta is higher during approach to walnut (green), and lower during walnut eating (blue) (top). Bottom: same as top, but for recordings from the same mouse in the cricket assay. e Left: Average activity in the l/vlPAG vgat projection to zona incerta during walnut approach and eating. Right: same as left, but for the cricket assay. The l/vlPAG vgat projection to zona incerta is activated prior to approach (f) and is inhibited following eating onset (g) in both assays (walnut n = 5, cricket n = 8; one-sample t-test, approach t-statistics: walnut=3.03, p = 0.039, cricket = 4.37, p = 0.003; eat t-statistics: walnut = −4.73, p = 0.009, cricket = −6.85, p < 0.001). h Mice were exposed to the environment depicted in the scheme. Mice were able to see and smell the walnut through the wire mesh but were not able to eat or touch it. i Heat map showing approach activity in the l/vlPAG vgat projection to the zona incerta for the assay shown in h. Note activity is highest in the entrance to the walnut corridor. j Average activity during approach onset to object or walnut. k Significantly elevated activity was observed during approach to walnut, but not object (n = 5; one-sample t-test, t-statistic (walnut) = 3.45, p = 0.026). ***p < 0.001, **p < 0.01, *p < 0.05. Data are presented as mean values +/- SEM. All error bars and error bands correspond to +/- SEM. Source data are provided as a Source Data File.
In order to test the necessity of the l/vlPAG vgat projection to ZI for appetitive drive, we expressed optogenetically inhibited Arch-expressing axon terminals of this projection in the walnut and cricket assays (Fig. 8a). Remarkably, inhibition of this projection decreased the amount of walnut consumed (Fig. 8c), increased the latency to first cricket attack and decreased the probability of hunting a cricket (Fig. 8d). We next tested if activity in this projection was sufficient to increase these behaviors. We thus expressed the excitatory opsin ChR2 in l/vlPAG vgat cells and implanted fiberoptic cannulae over the ZI (Fig. 8a). Optogenetic activation of this projection with delivery of blue light increased walnut consumption (Fig. 8e), decreased the latency to cricket predation and increased the probability of killing the cricket (Fig. 8f). It is possible that optogenetic terminal activations induce antidromic action potentials. However, our optogenetic projection inhibition of l/vlPAG vgat terminals in ZI decreased food-seeking, and these data are not confounded by potential antidromic effects (Fig. 8c, d). Our data thus indicate that higher activity in the l/vlPAG vgat projection to the ZI increases feeding. As the l/vlPAG vgat cells are GABAergic, it is expected that activation of this projection will inhibit the ZI. However, prior data are in disagreement with our results, as it has been shown that ZI inhibition decreases food-seeking7,8. To resolve this issue, we investigated the dynamics of this circuit. We expressed GCaMP6f in l/vlPAG vgat cells and ZI vgat cells contralaterally to avoid contamination of cell body signals with signals from long-range axon terminals. Fiber photometry recordings from contralateral hemispheres show that vgat cells from both the l/vlPAG and the ZI are activated during approach to food (Supplementary Fig. 17). These data suggest that the l/vlPAG vgat cells are not broadly inhibiting ZI vgat cells.

a Left: scheme showing viral strategy to express the opsins Arch (inhibitory) or ChR2 (excitatory) in l/vlPAG vgat cells. A fiberoptic cannula was placed over the zona incerta (ZI) to inhibit or excite the l/vlPAG vgat projection to the zona incerta. Right: Image showing ChR2-YFP expressing l/vlPAG axon terminals in the ZI. Opt: optic tract. Similar images were obtained in all 21 animals used. b Experimental timeline. c Inhibition of the l/vlPAG vgat projection to the zona incerta decreased eating of walnut (YFP n = 10, Arch n = 8; left: Wilcoxon rank-sum, z-value = −1.60, p = 0.11; right: Wilcoxon rank-sum, z-value = 2.45, p = 0.014; Wilcoxon signed rank (Arch), z-value = −2.52, p = 0.006). d This manipulation also increased latency to first cricket attack and decreased the probability of killing the cricket (YFP n = 13, Arch n = 8; left: Wilcoxon rank-sum, z-value = −2.03, p = 0.042, Wilcoxon signed rank (Arch), z-value = 2.12, p = 0.034; right: Wilcoxon rank-sum, z-value = 2.06, p = 0.039). e Excitation of the l/vlPAG vgat projection to the zona incerta increased consumption of walnut (YFP n = 6, ChR2 n = 9; left: Wilcoxon signed rank (ChR2), z-value = −2.52, p = 0.006; right: Wilcoxon rank-sum, z-value = −2.53, p = 0.006; Wilcoxon signed rank (ChR2), z-value = 2.52, p = 0.006). f This manipulation also decreased latency to successful cricket predation and increased the probability of killing the cricket (YFP n = 6, ChR2 n = 9; middle: Wilcoxon rank-sum, z-value = 3.18, p < 0.001, Wilcoxon signed rank (ChR2), z-value = −2.67, p = 0.008; right: Wilcoxon rank-sum, z-value = −2.47, p = 0.014; Wilcoxon signed rank (ChR2), z-value = 2.46, p = 0.014). ***p < 0.001, **p < 0.01, *p < 0.05, † p = 0.11. Data are presented as mean values +/- SEM. Source data are provided as a Source Data File.
To causally test this idea, we expressed the red-shifted opsin ChRmine3.3 in l/vlPAG vgat cells and the calcium indicator GCaMP6f in ZI vgat cells (Supplementary Fig. 18a, b). Remarkably, optogenetic excitation of l/vlPAG vgat cells in freely-moving mice increased neural activity in ZI vgat cells time-locked to laser ON epochs. The same manipulation had no effect in ZI vgat cells during isoflurane anesthesia. This result indicates that the increase in ZI vgat GCaMP6f fluorescence during laser ON epochs is due to increased ZI neural activity, and this effect was not caused by contamination by laser light artifacts (Supplementary Fig. 18c, d). Thus, both correlative (Supplementary Fig. 17) and perturbative data (Supplementary Fig. 18) show that the l/vlPAG input activates ZI vgat cells in freely-moving mice. These results reconcile our data with prior reports showing that higher ZI vgat activity causes increased food-seeking7,8. Taken together, our data show, activity in the l/vlPAG vgat to ZI projection is sufficient and necessary for appetitive drive of both non-prey and prey food types, showing that a bottom-up mechanism originating in the midbrain can potently affect food consumption.
Discussion
Actions related to exploration and investigation are of central importance, as these processes are the initial actions that lead to foraging, hunting and other behaviors. Despite their relevance, the mechanisms controlling such actions remain elusive. Here, we show that l/vlPAG vgat cells encode and bidirectionally control these behaviors, indicating a role in foraging (Fig. 3). Indeed, l/vlPAG vgat activity is sufficient and necessary for foraging and consumption. Optogenetically activating these cells produces foraging, exploration (Fig. 4b–h), and even induces aspects of compulsive eating (Fig. 4i). We also show that vgat l/vlPAG cells are bidirectionally connected with known feeding nodes, and that these cells control feeding via a projection to the ZI. Though a role for the l/vlPAG in mice hunting crickets has been described, feeding in most modern humans does not involve hunting and mostly consists of non-prey foods. Since the l/vlPAG vgat to ZI projection controls foraging and eating of both prey and non-prey foods (Fig. 8), it may be relevant for feeding in humans as well. The data suggest the l/vlPAG may have a broad role in feeding, rather than a species-specific role in cricket hunting in mice. Lastly, though several higher order inputs to the PAG have been shown to control hunting7,9,11,12,14, here we identify a bottom-up foraging and consumption circuit originating in the PAG.
Individual vgat l/vlPAG neurons show conserved eating-related modulation across different feeding assays
We characterized PAG vgat activity during appetitive and consummatory actions induced both by prey and non-prey food. These cells displayed increased activity preceding consumption (Fig. 2c, d) and prominent decreased activity following eating onset (Fig. 2e, f). This pattern was conserved across both feeding assays tested (Fig. 2k, m). Interestingly, cells that were inhibited during consumption showed correlated activity profiles across cricket and walnut assays (Fig. 2m). In contrast, cells activated by consumption were uncorrelated across assays (Fig. 2m). These data show negative modulation of vgat PAG cells during eating (Fig. 2m) is the most conserved pattern during the consummatory phase, regardless of food type.
PAG vgat neurons induce context-specific foraging actions
Environmental cues interact directly with the motivation to promote behavioral adaptation. Accordingly, we have shown previously that other PAG circuits function to integrate multisensory signals25 and encode approach or avoidance states26. Our data showed that vgat PAG neurons were mostly activated in the moments that precede eating in different situations (Fig. 2c, d). Indeed, a diversity of behavioral assays suggest that the activation of these neurons is associated with increased exploration (Fig. 4b–d), appetitive effects (Fig. 3e, f) and positive motivation (Fig. 4e,f,i). These effects were context-dependent, as unique behaviors were displayed in different contexts during PAG vgat activation (Fig. 4). Furthermore, activating these cells also elicited a strong motivation to pursue and follow objects (Fig. 4f). Here we show that these targeted-directed actions towards objects are flexible and depend on the resources available in the environment. In the absence of food, vgat l/vlPAG activation creates exploration and engagement with objects, but in the presence of food the directed actions preferentially target food (Fig. 4h).
Activation of l/vlPAG vgat cells does not induce locomotion per se, but rather elicits exploration of the available stimuli of interest, which may be holes in the holeboard task, an object or a hanging walnut in the other task (Fig. 4). An increase in speed during optogenetic stimulation was only observed when the exploratory action required higher locomotion (Supplementary Fig. 12). Consequently, optogenetic stimulation did not alter speed in the empty open field (an environment that does not have anything to explore - Supplementary Fig. 7c) or in the hanging walnut assay (where exploration of the walnut did not require increased movement - Supplementary Fig. 12). Similarly, endogenous l/vlPAG vgat activity also did not correlate with speed, and their neural activity could not be used to decode speed (Supplementary Fig. 7b). Taken together, these data show that l/vlPAG activity neither controls nor correlates with overall locomotion. In contrast, it is critical to point out that inhibition of feeding-inducing dorsal raphe vgat cells increases locomotion speed27,28. As l/vlPAG vgat cells do not directly affect overall locomotion, our data indicate that the observed results in our work were not due to optogenetic activation of raphe vgat cells. Accordingly, viral expression did not reach the dorsal raphe (Supplementary Fig. 9).
Activation of l/vlPAG vgat neurons also induced perseverative eating in the presence of an aversive stimuli indicative of involvement of this neural population in a strong motivational process potentially related to compulsive eating behaviors (Fig. 4i). Furthermore, inhibition of these neurons impaired both walnut consumption and reduced the probability of predation (Fig. 3d-c). These effects are not related to a decrease in fear or anxiety, as no correlation was observed between neural activity and threat proximity, and neither optogenetic activation nor inhibition impacted approach to threat (Supplementary Fig. 15), revealing the presence of PAG circuits that are relatively specialized for the pursuit of caloric resources.
Intriguingly, activation of the projection from lateral hypothalamus to PAG induces both hunting and attacking of conspecifics concomitantly with real-time place aversion14. In contrast, activation of PAG vgat cells induces foraging and real-time place preference (Fig. 4), but not aggression towards other mice (Supplementary Fig. 11), indicating these cells selectively control foraging-related behaviors. These results suggest that vgat PAG neurons produce positive motivation associated with driving appetitive responses.
PAG vgat neurons are bidirectionally connected to a food-seeking network
Here, we describe a bottom-up ascending pathway mediated by vgat PAG neurons to control appetitive and consummatory behaviors. Though the PAG has been viewed as a downstream target of both approach7,9,11,12,14 and avoidance behaviors22,29,30,31,Elevated plus maze The arms of the elevated plus maze were 30 × 7 cm. The height of the closed arm walls were 20 cm high. The maze is elevated 65 cm from the floor and is placed in the center of the behavior room away from other stimuli. Mice were placed in the center of the elevated plus maze facing a closed arm. For optogenetic experiments, light was delivered in alternating 2-min epochs for five epochs, totalling 10-min exposure. We used a long rectangular chamber (70 × 25 × 30 cm) as previously described25. Mice were acclimated to this environment for at least two days for 10 min each day. During rat exposure, a live rat is restrained to one end of the chamber using a harness attached to a cable with one end taped to the chamber wall. For the optogenetic experiments light alternated off or on in 3-min epochs for 4 epochs, totalling in a 12-min trial. The mice were habituated to a U-shaped maze enclosed arena (dimension: 24 × 20 × 13 cm) for 10 min on the day before the fiber photometry experiment. Each corridor (8 cm wide) was separated by an 18 cm wall, and had a window situated on its end. Then, a plastic cap (3 cm diameter) or a piece of walnut was positioned at the end and outside of the window of each corridor. The interaction with the object or the food was limited by a wired mesh positioned at the window. Mouse approaches to the windows were recorded from a camera situated at the top of the maze. The mean df/F activity during approach to object or walnut was used for the quantitative analyses. Mice were placed in a small chamber of dimensions 29 × 19 ×13 cm for 8 min in the presence of a novel conspecific of the same sex. Baseline was recorded for 5 min and optogenetic stimulation was performed alternating 1-min epochs for three epochs. Photometry was performed as described in detail previously54. Briefly, we used a 405-nm LED and a 470-nm LED (Thorlabs, M405F1 and M470F1) for the Ca2+-dependent and Ca2+-independent isosbestic control measurements. The two LEDs were band-pass filtered (Thorlabs, FB410-10 and FB470-10) and then combined with a 425-nm long-pass dichroic mirror (Thorlabs, DMLP425R) and coupled into the microscope using a 495-nm long-pass dichroic mirror (Semrock, FF495-Di02-25 3 36). Mice were connected with a branched patch cord (400 mm, Doric Lenses, Quebec, Canada) using a zirconia sleeve to the optical system. The signal was captured at 20 Hz (alternating 405-nm LED and 470-nm LED). To correct for signal artifacts of a non-biological origin (i.e., photo-bleaching and movement artifacts), custom Matlab scripts leveraged the reference signal (405-nm), unaffected by calcium saturation, to isolate and remove these effects from the calcium signal (470-nm). Correlation of df/F activity and threat proximity during EPM test and Rat assay were calculated as previously described26. To investigate the effects of l/vlPAG vgat stimulation in ZI vgat neural correlates, we optogenetically stimulated (20 Hz, 5 ms pulses, 2-minute light ON epochs) l/vlPAG vgat cells using ChRmine, an excitatory red-shifted opsin while recording ZI vgat activity. Mice were habituated to the test chamber for 10 min on the day prior to the test. On the test day, mice were introduced to an empty chamber without food, and ZI activity was recorded during l/vlPAG optogenetic stimulation. Each photostimulation consisted of 2-min epochs and were presented five times. Average activity in ZI vgat cells during light ON and light off epochs was calculated by averaging the mean signal for all light ON and light off epochs separately for each mouse, with a total average for both conditions calculated from all 4 mice. Epochs were identified using timestamp files defining laser onset and laser offset for all mice. On-off calculation is the difference in mean df/f (average light ON df/f minus the average light off df/f). All videos were recorded at 30 frames/sec using a Logitech HD C310 webcam and custom-built head-mounted UCLA miniscope. Open-source UCLA Miniscope software and hardware (http://miniscope.org/) were used to capture and synchronize the neural and behavioral videos25. The open-source UCLA miniscope analysis package (https://github.com/daharoni/Miniscope_Analysis) was used to motion correct the miniscope videos55. They were then temporally downsampled by a factor of four and spatially downsampled by a factor of two. The cell activity and footprints were extracted using the open-source package Constrained Nonnegative Matrix Factorization for microEndoscopic data (CNMF-E; https://github.com/zhoupc/CNMF_E)56. Only cells whose variance was greater than or equal to 10% of the maximum variance among non-outliers were used in the analysis25. Neurons were coregistered across sessions using the open-source probabilistic modeling package CellReg (https://github.com/zivlab/CellReg)57. To extract the pose of freely-behaving animals in the described assays, we implemented DeepLabCut58, an open-source convolutional neural network-based toolbox, to identify mouse nose, ear and tailbase xy-coordinates in each recorded video frame, as well as, when applicable, cricket xy-coordinates. These coordinates were then used to calculate speed and position at each time point. Approach was defined as movement epochs for which the mouse (1) had continuous movement towards the food source, (2) the distance towards the food source (either walnut or cricket) decreased by at least 10 cm and (3) the movement maintained a minimum velocity of 2 cm/s at all times. Eating was defined in timepoints in which the mouse is visibly biting or swallowing food. ‘Rearing’ was defined as epochs for which the mouse extended its body up the enclosure wall of either the walnut or cricket assay. This was automatically detected as samples when the mouse head position exceeded a minimum distance from either (1) the center of the circular cricket assay enclosure or (2) a minimum vertical threshold for the walnut eating assay. The DeepLabCut coordinates were not conducive to automatic classification of ‘cricket eating’, ‘walnut approach’, or ‘walnut eating’ behaviors. These behaviors were therefore scored manually by the experimenters. For individual cricket or walnut sessions, the multidimensional neural activity from each session was reduced to its top three principal components using Matlab function ‘pca’. For combined cricket/walnut session analysis, the session pairs were first coregistered using CellReg57, and the cell activity of putative neurons identified across sessions was first concatenated. The samples that corresponded with ‘approach’, ‘eat’, or ‘rear’ were plotted in 3-dimensional space. The silhouette score was calculated across these three behavioral clusters using the Matlab function ‘silhouette’. The Mahalanobis distance between cricket/walnut ‘approach’ and cricket/walnut ‘eat’ was calculated using Matlab function ‘mahal’. The pairwise Euclidean distance between the dimensionally-reduced ‘approach’ and ‘eat’ samples and their mean for concatenated cricket/walnut sessions was calculated using the Matlab function ‘pdist’. Discrete classification of behaviors was performed using multinomial logistic regression. Time-points following approach or eat by 2 s were selected, and a matched number of non-behavior time-points were randomly selected for training and validation. Each time point was treated as an individual data point. Training and validation were performed using five-fold cross- validation, with a minimum of 10 s between training and validation sets. As equal numbers of behavior and non-behavior samples were used to build the training and validation sets, chance accuracy was 50%. Sessions with less than five behaviors were excluded from the analysis. We used a Generalized Linear Model (GLM) to identify cells that showed increased calcium activity during each behavior. We fit this model to each cell’s activity, with behavior indices as the predictor variable and behavior coefficients as the measure of fit. Behavior onset times were then randomized 100 times and a bootstrap distribution built from the resulting GLM coefficients. A cell was considered a behavior-categorized cell if its coefficient was either greater than (+ cell) or less than (- cell) 95% of the bootstrap coefficient values. Coronal slices were prepared from vgat-Cre mice. Briefly, animals were deeply anesthetized with isoflurane and decapitated with sharp shears. The brains were placed and sliced in ice-cold modified artificial CSF (aCSF) containing the following: 194 mM sucrose, 30 mM NaCl, 4.5 mM KCl, 1 mM MgCl2, 26 mM NaHCO3, 1.2 mM NaH2PO4, and 10 mM D-glucose, saturated with 95% O2 and 5% CO2. A vibratome (DSK Microslicer; Ted Pella, Inc.) was used to cut 300 µm brain sections. The slices were allowed to equilibrate for 30 min at 33 °C in normal aCSF containing: 124 mM NaCl, 4.5 mM KCl, 2 mM CaCl2 1 mM MgCl2, 26 mM NaHCO3, 1.2 mM NaH2PO4, and 10 mM D-glucose continuously bubbled with 95% O2 and 5% CO2. Slices were then stored at 21–23 °C in the same buffer until use. All slices were used within 6 h of slicing. Procedures have been described in detail59,60. Electrophysiological recordings were performed as described previously for striatal slices59,61,62,63,64. The brain slice recordings were performed at RT (21–23 °C). Whole-cell patch-clamp recordings were made from neurons in the zona incerta using patch pipettes with a typical resistance of 3–5 MΩ. Neurons were morphologically and electrophysiologically identified. The intracellular solution for neurons membrane properties and evoked comprised the following:135 mM potassium gluconate, 3 mM KCl, 10 mM HEPES, 1 mM EGTA, 0.3 mM Na-ATP, 4 mM Mg-ATP, 0.1 mM CaCl2, 8 mM Na2-phosphocreatine, with pH adjusted to 7.3. To evoke IPSCs specifically from the l/vlPAG vgat cells, AAV5-EF1a-DIO-ChR2(H134R)-eYFP-WPRE-HGHpA was injected in the l/vlPAG and afference in the zona incerta were optogenetically stimulated with an LED light source (Lambda TLED + , sutter instrument) at 2 mW for 5 ms. Neurons were voltage-clamped at −60 mV and pre-incubated with drugs 5 min before recording. Evoked inhibitory postsynaptic currents were recorded in control condition, after incubation with 0.5 µM TTX (Tocris, #1069), 0.5 µM TTX + 100 µM 4-Aminopyridine (Tocris, #0940) or 25 µM bicuculline (Tocris, #0130). All recordings were performed at RT, using pCLAMP11.2 (Axon Instruments, Molecular Devices) and a MultiClamp 700B amplifier (Axon Instruments, Molecular Devices). Cells with access resistance that exceeded 20 MΩ were excluded from analysis. Analysis was performed using ClampFit 10.7 software. To calculate the mean peak amplitude for a cell on a given session, the local maxima were identified from the raw calcium activity (Matlab function ‘findpeaks’), and these maxima amplitudes were averaged. To identify the cell specificity of vgat and gad2 cells in the PAG, a new analysis of the data set from ref. 65 was performed using the same procedures as previously described. Nonparametric two-tailed Wilcoxon signed rank or rank-sum tests were used unless otherwise stated using MATLAB. Two-tailed tests were used throughout with α = 0.05. Asterisks in plots indicate the p values. The standard error of the mean was plotted in each figure as an estimate of variation. Multiple comparisons were adjusted with the false discovery rate method. All error bars in the plots represent the standard error of the mean. Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.Rat assay
U-shaped maze with walnut and object
Aggression
Fiber photometry
Simultaneous ZI vgat photometry recording and l/vl/PAG optogenetics
Miniscope video capture
Miniscope post-processing
Behavioral quantification
Behavioral classification
Principal component analysis and silhouette score
Behavior decoding
Behavior cell classification
Brain Slice Preparation and electrophysiological recordings in the zona incerta
Mean peak amplitude
Merfish data
Statistics
Reporting summary
Data availability
Source Data are provided with this paper. Raw miniscope data is available at the following link: https://datadryad.org/stash/share/cWBUCf87LWhzH9NTAfZwi26nEpWueEOd3drFnQdgXpA. Source data are provided with this paper.
Code availability
The associated code can be found at https://github.com/schuettepeter/vgat_dpag.
References
Yu, H. et al. Periaqueductal gray neurons encode the sequential motor program in hunting behavior of mice. Nat. Commun. 12, 1–15 (2021).
Comoli, E., Ribeiro-Barbosa, E. R., Negrão, N., Goto, M. & Canteras, N. S. Functional map** of the prosencephalic systems involved in organizing predatory behavior in rats. Neuroscience 130, 1055–1067 (2005).
Comoli, E., Ribeiro-Barbosa, E. R. & Canteras, N. S. Predatory hunting and exposure to a live predator induce opposite patterns of Fos immunoreactivity in the PAG. Behav. Brain Res. 138, 17–28 (2003).
Furigo, I. C. et al. The role of the superior colliculus in predatory hunting. Neuroscience 165, 1–15 (2010).
Ahmadlou, M. et al. A cell type–specific cortico-subcortical brain circuit for investigatory and novelty-seeking behavior. Science 372, eabe9681 (2021).
Ogasawara, T. et al. A primate temporal cortex-zona incerta pathway for novelty seeking. Nat. Neurosci. 25, 50–60 (2022).
Zhao, Z.-D. et al. Zona incerta GABAergic neurons integrate prey-related sensory signals and induce an appetitive drive to promote hunting. Nat. Neurosci. 22, 921–932 (2019).
Zhang, X. & van den Pol, A. N. Rapid binge-like eating and body weight gain driven by zona incerta GABA neuron activation. Science 356, 853–859 (2017).
Park, S.-G. et al. Medial preoptic circuit induces hunting-like actions to target objects and prey. Nat. Neurosci. 21, 364–372 (2018).
Hao, S. et al. The lateral hypothalamic and BNST GABAergic Projections to the anterior ventrolateral periaqueductal gray regulate feeding. Cell Rep. 28, 616–624.e5 (2019).
Han, W. et al. Integrated control of predatory hunting by the central nucleus of the Amygdala. Cell 168, 311–324.e18 (2017).
Roman-Ortiz, C., Guevara, J. A. & Clem, R. L. GABAergic basal forebrain projections to the periaqueductal gray promote food consumption, reward and predation. Sci. Rep. 11, 1–12 (2021).
Tryon, V. L. & Mizumori, S. J. Y. A novel role for the periaqueductal gray in consummatory behavior. Front. Behav. Neurosci. 12, 178 (2018).
Li, Y. et al. Hypothalamic circuits for predation and evasion. Neuron 97, 911–924.e5 (2018).
Motta, S. C., Carobrez, A. P. & Canteras, N. S. The periaqueductal gray and primal emotional processing critical to influence complex defensive responses, fear learning and reward seeking. Neurosci. Biobehav. Rev. 76, 39–47 (2017).
Yeh, L.-F., Ozawa, T. & Johansen, J. P. Functional organization of the midbrain periaqueductal gray for regulating aversive memory formation. Mol. Brain 14, 136 (2021).
Johansen, J. P., Tarpley, J. W., LeDoux, J. E. & Blair, H. T. Neural substrates for expectation-modulated fear learning in the amygdala and periaqueductal gray. Nat. Neurosci. 13, 979–986 (2010).
Kim, E. J. et al. Dorsal periaqueductal gray-amygdala pathway conveys both innate and learned fear responses in rats. Proc. Natl Acad. Sci. 110, 14795–14800 (2013).
Silva, C. & McNaughton, N. Are periaqueductal grey and dorsal raphe the foundation of appetitive and aversive control? A comprehensive review. Prog. Neurobiol. https://doi.org/10.1016/j.pneurobio.2019.02.001 (2019).
Marín-Blasco, I. J., Rangel, M. J., Baldo, M. V. C., Motta, S. C. & Canteras, N. S. The lateral periaqeductal gray and its role in controlling the opposite behavioral choices of predatory hunting and social defense. bioRxiv 2020.09.02.273961 https://doi.org/10.1101/2020.09.02.273961 (2020).
Krout, K. E. & Loewy, A. D. Periaqueductal gray matter projections to midline and intralaminar thalamic nuclei of the rat. J. Comp. Neurol. 424, 111–141 (2000).
Tovote, P. et al. Midbrain circuits for defensive behaviour. Nature 534, 206–212 (2016).
La-Vu, M. Q. et al. Sparse genetically defined neurons refine the canonical role of periaqueductal gray columnar organization. Elife 11, e77115 (2022).
Vianna, D. M., Graeff, F. G., Brandão, M. L. & Landeira-Fernandez, J. Defensive freezing evoked by electrical stimulation of the periaqueductal gray: comparison between dorsolateral and ventrolateral regions. Neuroreport 12, 4109–4112 (2001).
Reis, F. M. C. V. et al. Shared dorsal periaqueductal gray activation patterns during exposure to innate and conditioned threats. J. Neurosci. 41, 5399–5420 (2021).
Reis, F. M. et al. Dorsal periaqueductal gray ensembles represent approach and avoidance states. Elife 10, e64934 (2021).
Nectow, A. R. et al. Identification of a brainstem circuit controlling feeding. Cell 170, 429–442.e11 (2017).
Schneeberger, M. et al. Regulation of energy expenditure by Brainstem GABA neurons. Cell 178, 672–685.e12 (2019).
Wang, W. et al. Coordination of escape and spatial navigation circuits orchestrates versatile flight from threats. Neuron 109, 1848–1860.e8 (2021).
Wang, W. et al. Dorsal premammillary projection to periaqueductal gray controls escape vigor from innate and conditioned threats. Elife 10, e69178 (2021).
Evans, D. A. et al. A synaptic threshold mechanism for computing escape decisions. Nature 558, 590–594 (2018).
Deng, H., **ao, X. & Wang, Z. Periaqueductal gray neuronal activities underlie different aspects of defensive behaviors. J. Neurosci. 36, 7580–7588 (2016).
Hasue, R. H. & Shammah-Lagnado, S. J. Origin of the dopaminergic innervation of the central extended amygdala and accumbens shell: a combined retrograde tracing and immunohistochemical study in the rat. J. Comp. Neurol. 454, 15–33 (2002).
Kincheski, G. C., Mota-Ortiz, S. R., Pavesi, E., Canteras, N. S. & Carobrez, A. P. The dorsolateral periaqueductal gray and its role in mediating fear learning to life threatening events. PLoS One 7, e50361 (2012).
**e, Z. et al. Mechanically evoked defensive attack is controlled by GABAergic neurons in the anterior hypothalamic nucleus. Nat. Neurosci. 25, 72–85 (2022).
Rossier, D., Franca, V. L., Salemi, T., Natale, S. & Gross, C. T. A neural circuit for competing approach and defense underlying prey capture. Proc. Natl Acad. Sci. 118, e2013411118 (2021).
Chou, X.-L. et al. Inhibitory gain modulation of defense behaviors by zona incerta. Nat. Commun. 9, 1151 (2018).
Moffitt, J. R. et al. Molecular, spatial, and functional single-cell profiling of the hypothalamic preoptic region. Science 362, eaau5324 (2018).
Kosse, C., Schöne, C., Bracey, E. & Burdakov, D. Orexin-driven GAD65 network of the lateral hypothalamus sets physical activity in mice. Proc. Natl Acad. Sci. USA 114, 4525–4530 (2017).
Li, Z., Rizzi, G. & Tan, K. R. Zona incerta subpopulations differentially encode and modulate anxiety. Sci. Adv. 7, eabf6709 (2021).
Moon, H. C. & Park, Y. S. Reduced GABAergic neuronal activity in zona incerta causes neuropathic pain in a rat sciatic nerve chronic constriction injury model. J. Pain. Res. 10, 1125–1134 (2017).
Moon, H. C., Lee, Y. J., Cho, C. B. & Park, Y. S. Suppressed GABAergic signaling in the zona incerta causes neuropathic pain in a thoracic hemisection spinal cord injury rat model. Neurosci. Lett. 632, 55–61 (2016).
Jennings, J. H. et al. Visualizing hypothalamic network dynamics for appetitive and consummatory behaviors. Cell 160, 516–527 (2015).
Marino, R. A. M. et al. Control of food approach and eating by a GABAergic projection from lateral hypothalamus to dorsal pons. Proc. Natl Acad. Sci. 117, 8611–8615 (2020).
Carus-Cadavieco, M. et al. Gamma oscillations organize top-down signalling to hypothalamus and enable food seeking. Nature 542, 232–236 (2017).
Ye, Q., Nunez, J. & Zhang, X. Zona incerta dopamine neurons encode motivational vigor in food seeking. Science Advances 9, eadi5326 (2023).
Liu, K. et al. Lhx6-positive GABA-releasing neurons of the zona incerta promote sleep. Nature 548, 582–587 (2017).
Venkataraman, A. et al. Modulation of fear generalization by the zona incerta. Proc. Natl Acad. Sci. USA 116, 9072–9077 (2019).
Kolmac, C. I., Power, B. D. & Mitrofanis, J. Patterns of connections between zona incerta and brainstem in rats. J. Comp. Neurol. 396, 544–555 (1998).
Kolmac, C. & Mitrofanis, J. Distribution of various neurochemicals within the zona incerta: an immunocytochemical and histochemical study. Anat. Embryol. 199, 265–280 (1999).
Luo, L. Architectures of neuronal circuits. Science 373, eabg7285 (2021).
Fratzl, A. & Hofer, S. B. The caudal prethalamus: Inhibitory switchboard for behavioral control? Neuron 110, 2728–2742 (2022).
Nieh, E. H. et al. Decoding neural circuits that control compulsive sucrose seeking. Cell 160, 528–541 (2015).
Kim, C. K. et al. Simultaneous fast measurement of circuit dynamics at multiple sites across the mammalian brain. Nat. Methods 13, 325–328 (2016).
Aharoni, D. & Hoogland, T. M. Circuit investigations with open-source miniaturized microscopes: past, present and future. Front. Cell. Neurosci. 13, 141 (2019).
Zhou, P. et al. Efficient and accurate extraction of in vivo calcium signals from microendoscopic video data. Elife 7, e28728 (2018).
Sheintuch, L. et al. Tracking the same neurons across multiple days in Ca2+ imaging data. Cell Rep. 21, 1102–1115 (2017).
Mathis, A. et al. DeepLabCut: markerless pose estimation of user-defined body parts with deep learning. Nat. Neurosci. 21, 1281–1289 (2018).
Jiang, R., Diaz-Castro, B., Looger, L. L. & Khakh, B. S. Dysfunctional calcium and glutamate signaling in striatal astrocytes from Huntington’s Disease Model Mice. J. Neurosci. 36, 3453–3470 (2016).
Tong, X. et al. Astrocyte Kir4.1 ion channel deficits contribute to neuronal dysfunction in Huntington’s disease model mice. Nat. Neurosci. 17, 694–703 (2014).
Chai, H. et al. Neural circuit-specialized astrocytes: transcriptomic, proteomic, morphological, and functional evidence. Neuron 95, 531–549.e9 (2017).
Octeau, J. C. et al. An optical neuron-astrocyte proximity assay at synaptic distance scales. Neuron 98, 49–66.e9 (2018).
Srinivasan, R. et al. Ca(2+) signaling in astrocytes from Ip3r2(-/-) mice in brain slices and during startle responses in vivo. Nat. Neurosci. 18, 708–717 (2015).
Srinivasan, R. et al. New transgenic mouse lines for selectively targeting astrocytes and studying calcium signals in astrocyte processes in situ and in vivo. Neuron 92, 1181–1195 (2016).
Vaughn, E., Eichhorn, S., Jung, W., Zhuang, X. & Dulac, C. Three-dimensional interrogation of cell types and instinctive behavior in the periaqueductal gray. bioRxiv 2022.06.27.497769 https://doi.org/10.1101/2022.06.27.497769 (2022).
Acknowledgements
This work was supported by the National Institute for Mental Health (R00 MH106649 and R01 MH119089) (A.A.), the National Diabetes and Digestive and Kidney Diseases (R01DK139605 to A.A.), the Brain & Behavior Research Foundation (Grants # 29204 and # 22663 respectively to J.C.K., A.A., and # 27654, # 31093 to F.M.C.V.R.), the National Science Foundation (NSF-GRFP DGE-1650604, P.J.S.), the UCLA Affiliates fellowship (P.J.S.) and the Hellman Foundation (A.A.). We thank Judy Genshaft and Steven Greenbaum for funding the 2022 Young Investigator Award (Grant # 31093, F.M.C.V.R.) and The Nancy and Jon Glaser Family Friends of Semel Scholar Award (F.M.C.V.R.). A.T. was supported by F31MH127943-01A1. B.A.M. was supported by an NSF-GRFP award. JMI was supported with FAPESP grants #2016/17329-3 and #2019/17677-0. AHK was supported with grants from FAPESP #2019/17892-8 and CNPq #312047/2017-7. We thank Prof. C. Dulac for mentoring E. Vaughn and guiding the generation and analysis of the PAG MERFISH dataset.
Author information
Authors and Affiliations
Contributions
FMCVR performed behavioral experiments, surgeries, experimental and analysis design, conceptualized the project and wrote the paper. SMP performed behavioral experiments and data analysis. PJS performed experimental analysis and made the figures. MO and BK performed the electrophysiology experiments and analyzed the data. EV analyzed the MERFISH data. ES performed behavioral experiments and histology. DCTN, FTHY and EI performed histology. MS, JMI and AT assisted in behavioral and viral tracing experiments. BK, AJS, AHK and JCK provided mentoring for students and advice on data analysis. MC performed behavioral experiments. MS and AJS assisted with rabies tracing. BAM performed data analysis on dual site photometry and optogenetics-related recordings. AA wrote the paper and guided experimental and analysis design.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Wenfei Han and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Reis, F.M.C.V., Maesta-Pereira, S., Ollivier, M. et al. Control of feeding by a bottom-up midbrain-subthalamic pathway. Nat Commun 15, 2111 (2024). https://doi.org/10.1038/s41467-024-46430-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-46430-5
- Springer Nature Limited