Abstract
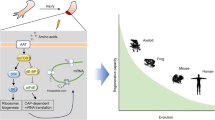
Regenerative capacity declines throughout evolution and with age. In this study, we asked whether metabolic programs underlying regenerative capability might be conserved across species, and if so, whether such metabolic drivers might be harnessed to promote tissue repair. To this end, we conducted metabolomic analyses in two vertebrate organ regeneration models: the axolotl limb blastema and antler stem cells. To further reveal why young individuals have higher regenerative capacity than the elderly, we also constructed metabolic profiles for primate juvenile and aged tissues, as well as young and aged human stem cells. In joint analyses, we uncovered that active pyrimidine metabolism and fatty acid metabolism correlated with higher regenerative capacity. Furthermore, we identified a set of regeneration-related metabolite effectors conserved across species. One such metabolite is uridine, a pyrimidine nucleoside, which can rejuvenate aged human stem cells and promote regeneration of various tissues in vivo. These observations will open new avenues for metabolic intervention in tissue repair and regeneration.
Similar content being viewed by others

Introduction
Regeneration is the process of rejuvenating or replacing damaged, diseased, or aged tissues1. From lower animals to humans, every species is endowed with a certain degree of regeneration. For example, axolotl, the Mexican salamander, or the “walking fish”, is evolutionarily primitive vertebrate known to possess a higher regenerative capacity than mammals2,3,4,5. Another example is the deer antler, which is the only organ capable of complete regeneration in mammals6,7,8. In most mammals, the limited anatomical and functional recovery capabilities reside in young tissue and decline with age2,9,10,11, leading to compromised tissue repair after injury.
Across species, stem cells usually take center stage in tissue repair and regeneration1,12. The axolotl can regenerate their limbs through the formation of blastema tissue, a mass of dedifferentiated stem cells3,5,13. Similarly, during annual regeneration, deer antler produces a whole organ containing blood vessels, cartilage, bone, dermis, and nerves from deer antler stem cells (dASCs) that express classic mesenchymal stem cell (MSC) markers and reside in the mid-beam antler periosteum7,14,15,16. Compared to stem cells from regenerative tissues of the axolotl limb and the deer antler, human stem cells, such as human mesenchymal stem cells (hMSCs), possess a relatively limited capacity for regenerative repair of damages to vital tissues and organs2,12, but gradually lose such capacity with age. Whether molecular characteristics between these naturally occurring regeneration processes are evolutionarily conserved across species is unknown.
Using comparative methods to describe the similarities and differences between species is a powerful strategy to discover the regulatory mechanisms that underline vital life events, such as regeneration. The effectiveness of this method depends on whether there are comparable and sufficient overlap** factors across different samples. Unlike proteins that are biomacromolecules, the structure of metabolites is relatively similar between species, making metabolism an ideal research area for investigating evolutionarily conserved biology17. Yet, a systematic and high-resolution metabolomic characterization across paradigms with high regenerative capacity in different species and different tissues has not yet been attempted. The rapid development of untargeted metabolomic profiling could serve this purpose18.
Here, we sought to understand how metabolic regulation intersects with inherent regenerative capacity using comparative approaches. Samples for this study included i) species that are more primitive on the evolutionary scale but can renew entire organs, and ii) higher species in evolution that have lost full organ regenerative capacity but retain a limited capacity for tissue repair. We systematically depicted metabolic profiles in various regeneration-related contexts, and we discovered that high pyrimidine and fatty acid metabolism was shared across species, tissues, and cells with high regenerative capacity. We identified uridine as a pro-regenerative metabolite that promoted human stem cell activity and enhanced regeneration in multiple tissues in mammals.
Results
Transcriptomic analysis revealing convergent metabolic pathways in models with enhanced regenerative potentials
We hypothesized that comparative studies of highly divergent regenerative models might reveal evolutionarily conserved programs driving tissue regeneration2,19. To test this hypothesis, we decided to profile the transcriptomes and metabolomes of samples from naturally occurring regeneration processes in vertebrate organ regeneration, and young and old primate tissues and human stem cell models with differential regenerative capacities20 (Fig. 1a). To represent whole organ regeneration, we chose the axolotl limb and deer antler. We obtained axolotl blastema (AB) at the amputation surface at day 11 post amputation (DPA 11), the timepoint when axolotl blastema stem cells (aBSCs) peak21 (Fig. 1a). Similarly, we isolated dASCs from the amputation surface. dASCs have a higher self-renewal ability than human primary mesenchymal stem cells (hPMSCs)6,7,16 (Fig. 1a and Supplementary Fig. S1a–c). In human stem cell model, young wild-type hMSCs manifested higher self-renewal and regenerative abilities relative to prematurely aged control (WRN-depleted hMSCs, mimicking Werner syndrome (WS), a human progeroid syndrome)22,23,24,25,26,27 (Fig. 1a and Supplementary Fig. S1d–f). To represent young tissues, we obtained eight tissues/organs from young and old non-human primates (NHPs) for comparison. The tissues (i.e., liver, skeletal muscle, skin, kidney, brain, heart, white adipose tissue (WAT), as well as blood plasma) selected for further analysis also hold varied regenerative abilities (Fig. 1a).

a Flowchart of experimental design for obtaining transcriptome and metabolome data from samples with differential regenerative capacities: axolotl blastema (AB) at DPA 0 and DPA 11, deer antler stem cells (dASCs), tissues from young and aged non-human primates (NHPs), and young and aged hMSCs. b Left, PCA analysis of the transcriptome data of AB at DPA 0 and DPA 11 (top) and WT and WS hMSCs (bottom). Right, tree plot showing the Euclidean distance for transcriptome data of young and aged NHP tissues. c Circos plots showing the overlap of upregulated (top) and downregulated (bottom) DEGs in AB at DPA 11, young tissues, and young hMSCs. d Bubble plot showing the enriched GO terms and pathways for upregulated DEGs in AB at DPA 11, young tissues, and young hMSCs. The color key from white to amaranth indicates low to high –log10(P-value). e Left, bubble plot showing the relative differential expression (DE) score for all genes and regeneration genes in AB at DPA 11, young tissues, and young hMSCs. The color key from white to amaranth indicates the relative DE scores from low to high. Right, bubble plot showing the convergently upregulated regeneration DEGs in AB at DPA 11, young tissues, and young hMSCs. The color key from white to amaranth indicates log2(fold change) values of DEGs from low to high. Genes convergently upregulated in at least four tissues/cells with high regenerative capacity were shown. f Left, bubble plot showing the relative DE score for all genes, mitochondria-related genes or metabolic genes in AB at DPA 11, young tissues, and young hMSCs. Middle, bubble plot showing the convergently upregulated mitochondria-related DEGs in AB at DPA 11, young tissues, and young hMSCs. Right, bubble plot showing the convergently upregulated metabolic DEGs in AB at DPA 11, young tissues, and young hMSCs. The color key from white to amaranth indicates DE scores (left) or log2(fold change) values of DEGs (middle and right) from low to high. Genes convergently upregulated in at least four tissues/cells with higher regenerative capacity were shown.
Genome-wide RNA sequencing analysis revealed that differentially expressed genes (DEGs) overlapped extensively across axolotl and NHP tissues, and genes involved in regeneration-related Gene Ontology (GO) terms or pathways, including “response to growth factor” and “tissue morphogenesis” were upregulated in axolotl blastema at DPA 11 and young NHP tissues (Fig. 1b–d and Supplementary Fig. S1g–h). Interestingly, the upregulated DEGs were convergently enriched in metabolism-related terms, including “mitochondrial organization”, “generation of precursor metabolites and energy” and “nucleotide metabolic process” (Fig. 1d and Supplementary Fig. S1h).
As expected, convergently upregulated DEGs, included known regeneration-related genes, such as growth factors or transcriptional factors required for organismal development (Fig. 1e). Of note, PPARGC1A, the master gene of mitochondrial biogenesis and fatty acid metabolism, was upregulated in most of the young tissues (Fig. 1e). Concomitantly, we also observed convergent upregulation of mitochondrial and metabolic pathways in tissues/cells with higher regenerative capacity (Fig. 1f). Among them, ACADVL, ACADS, ECH1, and ECI1 are all enzymes that catalyze fatty acid oxidation (FAO) in mitochondria (Fig. 1f). For example, ACADVL, the dehydrogenase that catalyzes the first step of mitochondrial FAO28,29, was upregulated in most of the young tissues (Fig. 1f). These results imply that metabolic regulation is an important feature associated with regenerative potential, and suggest that our framework approach may unveil metabolic commonalities across species and tissues.
Metabolomic analysis unveiling conservation and difference in metabolic characteristics between species
Metabolic processes are fundamental for organismal growth and development, and are required for the coordinated regulation of regeneration30,31. Indeed, we found that the convergent changes of metabolism-related genes across species were more prominent than that of global gene expression between samples with differential regeneration capacities (Fig. 1f). Next, we employed ultrahigh performance liquid chromatography-tandem mass spectroscopy (UPLC-MS/MS)-based metabolomics to profile metabolites across our models with differential regenerative abilities (Fig. 1a). After stringent quality control and normalization, we identified a range of 400–759 metabolites of various classes (including lipids, nucleotides, amino acids, carbohydrates, energy, peptides, cofactors, and vitamins) in each of the phylogenetically distant models (Fig. 2a and Supplementary Fig. S2a). However, we found that the metabolite distributions segregating with higher regenerative capacity were overall comparable across the axolotl blastema, dASCs, eight tissues of NHPs, and human stem cells (Fig. 2a), indicative of a considerable level of evolutionary conservation.

a Pie plot showing the percentage of super-pathway for identified metabolites in AB, dASCs, NHP tissues and hMSCs. b PLS-DA analysis of metabolomic data generated in AB, dASCs, NHP tissues and hMSCs. c Bar plot showing the count of DPMPs identified in AB at DPA 11, dASCs, young NHP tissues, and young hMSCs. The percentages of DPMPs in each super-pathway were shown on the top of each bar graph. d The hierarchical clustering dendrogram showing the similarity of the metabolic profile changes in AB at DPA 11, dASCs, young NHP tissues, and young hMSCs. e Heatmap showing the differential abundance (DA) for identified metabolites in each super-pathway in AB at DPA 11, dASCs, young NHP tissues, and young hMSCs. The color key from blue to amaranth indicates DA score from low to high for each super-pathway. f Heatmap showing the differential abundance for identified metabolites in each sub-pathway in AB at DPA 11, dASCs, young NHP tissues, and young hMSCs. Sub-pathways increased in at least seven tissues/cells with higher regenerative capacity were highlighted. The color key from blue to amaranth indicates DA score from low to high for each sub-pathway. g Network diagram showing the representative sub-pathways and relative abundance for DPMPs in each sub-pathway. The color of the edge from grey to amaranth indicates log2(fold change) from low to high. The node sizes are positively correlated to the edge counts for each node.
To characterize metabolic features in samples with higher regenerative potential, we performed partial least squares discriminant analysis (PLS-DA) (Fig. 2b). Overall, our analysis showed a clear separation of metabolomes between samples with higher regenerative abilities and their control counterparts (Fig. 2b and Supplementary Fig. S2b), indicating a clear correlation between metabolic features and regenerative capacities. Next, we assessed the changes of metabolite abundance underlying differential regenerative potentials and identified differentially present metabolic products (DPMPs) (Fig. 2c). Globally, metabolic profiles were most influenced by species diversity, followed by tissue specificity (Fig. 2c, d). In addition, WAT had the most distinct metabolic changes out of all the tested tissues between old and young individuals, highlighting the uniqueness of WAT metabolism (Fig. 2c, d and Supplementary Fig. S2b). In all, these analyses demonstrate that metabolomic profiles of samples with higher regenerative potentials are clearly different from their less regenerative counterparts.
Identification of key metabolites associated with higher regenerative potential
Next, we assigned DPMPs to super-pathways and sub-pathways according to metabolite annotations (Fig. 2e, f). At the super-pathway level, lipids, amino acids, and nucleotides account for ~60% of metabolic changes in all models (Fig. 2c, e). In general, nucleotide metabolism appeared more prominent in the blastema and young NHP tissues, while lipid metabolism was highly abundant in almost all young NHP tissues and young hMSCs (Fig. 2e).
Within the super-pathways, we were intrigued to see a metabolite subset uniformly more abundant in almost all samples with higher regenerative potential than their regeneration-refractory counterparts (Fig. 2f). For example, uracil-containing pyrimidine metabolites in the nucleotide super-pathway were enriched in the blastema, dASCs, young WT-hMSCs, and almost all young NHP samples (Fig. 2f, g). Guanine-containing purine metabolites were abundant in the blastema, young WT-hMSCs, young NHP plasma, WAT, and liver (Fig. 2f). Pyrimidine and purine metabolism offer structural blocks for DNA and RNA synthesis, critical to vital biological processes, including development32. Metabolites in lipid metabolism sub-pathways, such as fatty acid (dicarboxylate) and lysophospholipid, were predominantly enriched in almost all samples (Fig. 2f, g, and Supplementary Fig. S2c), reflecting the metabolic potential of lipolysis in models with higher regenerative capacity. In addition, long-chain mono-unsaturated/saturated fatty acids, phosphatidylcholine (PC), and phosphatidylethanolamine (PE) were also substantially enriched in most of the young tissues or stem cells (Fig. 2f).
Four sub-pathways related to amino acid metabolism, namely, polyamine metabolism, urea cycle/arginine and glutathione metabolism, glycine, serine and threonine metabolism, were also enriched (Fig. 2f, g). Among these, glutathione metabolism plays an important role in defensing against oxidant33,34, and was recently reported to be activated in liver regeneration35. In addition, polyamine metabolites (e.g., spermine and spermidine) are well-known pro-regenerative metabolites36,37,38. Taken together, these data imply a metabolic preference underlying high regenerative ability.
Transcriptional regulation of metabolic pathways related to regenerative capacity
To dissect the identified regulatory pathways, we leveraged the genome-wide transcriptomic analysis with a focus on metabolic genes and conducted an integrated pathway-level analysis of transcriptomic and metabolomic data with the MetaboAnalyst web tool39 (Supplementary Fig. S3a). This analysis revealed that within each super-pathway, specific sub-pathways were activated in almost all models, i.e., “pyrimidine metabolism” in the nucleotide super-pathway, “glycerophospholipid metabolism”, “glycerol metabolism” and “fatty acid degradation” in the lipid super-pathway, and “glutathione metabolism” in the amino acid super-pathway (Supplementary Fig. S3a). In pyrimidine metabolism, both DPMPs (uridine and 2′-deoxyuridine) and differentially expressed metabolic genes (DEMGs) (i.e., UCK2, UCKL1, and RRM1) were highly represented in samples with higher regenerative potentials (Supplementary Fig. S3b).
Transcriptional regulatory network analysis of DEMGs further underscored that the regulons underlying metabolic regulation overlap across models (Supplementary Fig. S3c, d). Notably, almost all PPAR-RXR complex members were identified as core transcription factors, namely PPARA, PPARD, PPARG, and their coactivator PPARGC1A, as well as the retinoid X receptors (RXRA and RXRG) (Supplementary Fig. S3c, d). As one of the most prominent regulatory systems for maintaining lipid and glucose homeostasis, the PPAR-RXR complex regulates a suite of genes involved in lipid and nucleotide metabolic processes40,41,42. Consistently, we found that the genes involved in the PPAR signaling pathway were activated, including those involved in lipid transport (APOA1, APOA2, APOC3, and APOA5) and fatty acid transport (ACSL4 and SLC27A1) (Supplementary Fig. S3e). Additionally, the molecular program for FAO was also more active in samples with higher regenerative potentials, including mitochondrial genes essential for the uptake of long-chain fatty acids (CPT1 and CPT2) (Supplementary Fig. S3e). Overall, the combined metabolomic and transcriptomic datasets revealed a strong co-occurrence between lipid and pyrimidine metabolism and identified the PPAR-RXR complex as a potential hub to be involved in the regulation of regeneration.
Screening of natural metabolites reinforcing hMSC activity
We next asked if it was possible to identify a metabolite that could boost the activity of human stem cells. To answer this question, we identified 29 candidate metabolites as upregulated DPMPs in at least four tissues or cells with higher regenerative ability from the nucleotide, amino acid, and lipid super-pathways (Fig. 3a and Supplementary Table S1). Among them, palmitoyl dihydrosphingomyelin (d18:0/16:0), lignoceroyl sphingomyelin (d18:1/24:0), sphingomyelin (d18:1/24:1, d18:2/24:0), and sphingosine in lipid super-pathway and uridine, 2′-deoxyuridine, 2′-O-methylguanine, and 7-methylguanine in nucleotide super-pathway were enriched in at least 4 tissues or cells with a higher regenerative ability (Fig. 3a).

a Bubble plot showing the increased metabolites in at least four tissues/cells with higher regenerative capacity. The bubble sizes are positively correlated to the log2(fold change) values. b Top-left, schematic illustration of the screening strategy for candidate metabolites reinforcing self-renewal activity of WS hMSCs. Top-right, scatter plot showing the relative cell proliferative abilities upon treatment with candidate metabolites at indicated concentrations in WS hMSCs. Bottom, top-ranked metabolites at indicated concentrations for cell proliferation are shown in the table. c Schematic representation (left) and quantitative data (right) of the detection of uridine concentration in the plasma of young (19–25 years old, n = 28) and aged (75–92 years old, n = 21) individuals. d Immunostaining of Ki67 in vehicle- and uridine (200 μM)-treated WS hMSCs (passage 5, P5) at P2 post treatment. Data are presented as the mean ± SEM (two-tailed unpaired Student’s t test). n = 3 biological replicates. Scale bars, 25 μm. e Cell cycle analysis of vehicle- and uridine (200 μM)-treated WS hMSCs (P5) at P2 post treatment. Data are presented as the mean ± SEM (two-tailed unpaired Student’s t test). n = 3 biological replicates. f SA-β-gal staining of vehicle- and uridine (200 μM)-treated WS hMSCs (P5) at P2 post treatment. Scale bars, 100 μm. Data are presented as the mean ± SEM (two-tailed unpaired Student’s t test). n = 3 biological replicates. g Toluidine blue staining analysis to evaluate the chondrogenesis of vehicle- and uridine (200 μM)-treated WS hMSCs (P5) at P2 post treatment. Data are presented as the mean ± SEM (two-tailed unpaired Student’s t test). n = 8 biological replicates. Scale bars, 100 μm. h Heatmap diagrams showing enriched GO terms and pathways for upregulated genes (left) and downregulated genes (right) in uridine (200 μM)-treated WS hMSCs (P5) at P2 post treatment as compared to vehicle-treated counterparts. The color keys from white to red or blue indicate the enrichment levels [–log10(P-value)] from low to high. i Gene set enrichment analysis (GSEA) showing representative GO terms and pathways in uridine (200 μM)-treated WS hMSCs (P5) at P2 post treatment as compared to vehicle-treated counterparts.
From this set of candidates, we screened commercially available metabolites (Supplementary Table S2) for their effects in promoting self-renewal of aged hMSCs (Fig. 3b), which is based on the known fact that regenerative ability in hMSCs is correlated with their self-renewal activity22,43,44. In line with the activated lipid metabolism in young hMSCs, sphingomyelin (d18:1/24:1, d18:2/24:0) supplementation stimulated hMSC self-renewal (Fig. 3a, b). More importantly, uridine, which we found to be more abundant in the plasma from young individuals than that from old individuals (Fig. 3c), was identified as a metabolite segregating with higher activity in WS hMSCs (Fig. 3b).
When we supplemented the culture medium with uridine, we found that uridine treatment was sufficient to reprogram the prematurely and physiologically aged stem cell models (WS/HGPS (Hutchinson-Gilford progeria syndrome)-hMSCs and hPMSCs) into a younger state with a higher regenerative ability (Fig. 3d–f and Supplementary Fig. S4a–f). Specifically, uridine-treated hMSCs achieved a much higher proliferation rate and an enhanced capacity to form cartilage and gained increased genome and epigenome stability (Fig. 3d–g and Supplementary Fig. S4a–f). In accordance, genome-wide RNA-seq analysis showed that upregulated genes were mainly associated with “cell cycle” and “DNA integrity checkpoint” GO terms or pathways (Fig. 3h). Consistent with a previous study reporting that uridine addition rescues pyrimidine biosynthesis deficiency45, we found that the “pyrimidine nucleoside metabolic process” was elevated by uridine supplementation (Fig. 3i). Uridine treatment also appears to have a beneficial role in mitochondrial activity, as we found augmented gene expression associated with “mitochondrial central dogma”, “mtDNA maintenance”, and “mitochondrial gene expression” in uridine-treated hMSCs (Fig. 3i). Taken together, these results showed that uridine supplementation drives broad transcriptional changes associated with improved hMSC activity.
Uridine treatment enhances regeneration and repair in various types of tissues
The extent of tissue repair after injury is limited by organismal intrinsic regenerative capacity46. Next, we asked whether uridine supplementation could promote regeneration or tissue repair in multiple tissues, including skeletal muscle, heart, liver, skin, and articular cartilage (Fig. 4a). Relative to vehicle-treated mice, we observed that uridine treatment promoted tissue repair in both muscular and cardiac injury models (Figs. 4b–j, 5a–f, and Supplementary Fig. S5a–d). For instance, uridine treatment facilitated muscle tissue regeneration, reduced fibrotic or erosion area, decreased proinflammatory cytokine levels, and endowed treated mice with higher grip strength and longer running distance (Fig. 4b–g). We next performed genome-wide RNA-seq analysis in injured muscles with or without uridine treatment (Fig. 4h–j). In line with the decreased levels of proinflammatory cytokines in mouse serum in the uridine-treated groups (Fig. 4e), bulk RNA sequencing showed uridine supplementation antagonized the expression of a panel of the inflammatory genes, the expression of which was elevated in injured muscles (Fig. 4j). In comparison, pathways related to muscle structure development, as well as metabolic pathways, especially in “small-molecule biosynthetic process” and “nucleotide metabolic process” were upregulated in uridine-treated mice (Fig. 4j). These data suggest that uridine supplementation, in turn, may promote regeneration and repair by remodeling metabolic adaptation. We next sought to dissect the cell type-specific effects associated with the regenerative response by constructing a single-nucleus transcriptomic atlas of uridine-treated muscle. We identified 14 muscle cell types, including satellite cells (Pax7+), the rare muscle stem cell population, fibro-adipogenic progenitors (FAPs, Pdgfra+), an interstitial mesenchymal cell population that supports muscle regeneration47, and fast-twitch muscle fibers (Mybpc2+ or Myh1+), that use anaerobic respiration to produce rapid movement bursts (Supplementary Fig. S5a, b). Similar to the bulk RNA-seq results, uridine supplementation restored the expression of genes associated with pyrimidine nucleotide biosynthesis and muscle structure development across cell types, especially in fast-twitch muscle fibers (Supplementary Fig. S5c).

a Schematic diagram showing the summarized phenotypes in tissue regeneration or repair models. b Schematic diagram for the time course of the mouse muscle cryoinjury and vehicle or uridine treatment. c Left, haematoxylin and eosin (H&E) staining of the skeletal muscle derived from sham mice (n = 10 mice) and mice treated with vehicle (n = 10 mice) or uridine (n = 10 mice) post cryoinjury. Right, quantitative data of mean myofiber cross-sectional area (CSA) in the skeletal muscle derived from sham mice and mice treated with vehicle or uridine post cryoinjury. Data are presented as the means ± SEM (two-tailed unpaired Student’s t-test). Scale bars, 100 μm. d Left, Masson staining of the quadriceps femoris derived from sham mice (n = 9 mice) and injured mice treated with vehicle (n = 9 mice) or uridine (n = 9 mice). Right, quantitative data of the fibrotic area. Data are presented as the means ± SEM (two-tailed unpaired Student’s t-test). Scale bars, 100 μm. e ELISA detecting the secretion of proinflammatory factors in the serum of sham mice (n = 20 mice) and mice treated with vehicle (n = 25 mice) or uridine (n = 25 mice) post cryoinjury. Data are presented as means ± SEM (two-tailed unpaired Student’s t-test). f Grip strength evaluation of the hind limbs of sham mice (n = 15 mice) and mice treated with vehicle (n = 15 mice) or uridine (n = 15 mice) at day 7 post cryoinjury. Data are presented as means ± SEM (two-tailed unpaired Student’s t test). g Treadmill distance of sham mice (n = 15 mice) and mice treated with vehicle (n = 15 mice) or uridine (n = 15 mice) at day 7 post cryoinjury. Data are presented as means ± SEM (two-tailed unpaired Student’s t-test). h Gene set enrichment analysis showing relative expression levels for downregulated DEGs upon cryoinjury (top) and upregulated DEGs upon cryoinjury (bottom) in the muscle tissues derived from vehicle- or uridine-treated mice. i Top, Venn diagram showing the downregulated genes upon cryoinjury and upregulated genes upon uridine treatment as compared to vehicle treatment. The overlapped genes were defined as “rescue DEGs (upregulated)” (left). Ring-heatmap plot showing the relative expression levels of “rescue DEGs (upregulated)” in mouse muscle regeneration model (right). Bottom, Venn diagram showing the upregulated genes upon cryoinjury and downregulated genes upon uridine treatment as compared to vehicle treatment. The overlapped genes were defined as “rescue DEGs (downregulated)” (left). Ring-heatmap plot showing the relative expression levels of “rescue DEGs (downregulated)” in mouse muscle regeneration model (right). The color key from blue to amaranth indicates log2(fold change) values from low to high. j Top, GO term and pathway enrichment analysis of “rescue DEGs (upregulated)” in mouse muscle regeneration model. Bottom, GO term and pathway enrichment analysis of “rescue DEGs (downregulated)” in mouse muscle regeneration model. The color keys from white to red or blue indicate –log10(P-value) from low to high.

a Schematic diagram for the time course of the mouse myocardial infraction (MI) modeling and vehicle or uridine treatment. Uridine or vehicle treatment were performed every other day, as indicated by the red arrows. b Left, representative echocardiographic images of sham mice (n = 6 mice) and mice treated with vehicle (n = 13 mice) or uridine (n = 13 mice) at day 7 after MI modeling and intramyocardial injection of vehicle or uridine. Right, quantitative data of left ventricular ejection fraction (LVEF) and left ventricular fractional shortening (LVFS). Data are presented as the means ± SEM (two-tailed unpaired Student’s t-test). c Quantitative analysis of LDH and CK level of sham mice (n = 15 mice) and mice treated with vehicle (n = 25 mice) or uridine (n = 25 mice) on the next day after MI modeling and intramyocardial injection of vehicle or uridine. Data are presented as the means ± SEM (two-tailed unpaired Student’s t-test). d Gene set enrichment analysis showing relative expression levels for downregulated DEGs upon myocardial infarction (top) and upregulated DEGs upon myocardial infarction (bottom) in the heart tissues from vehicle- or uridine-treated mice. e Top, Venn diagram showing the downregulated genes upon myocardial infarction and upregulated genes upon uridine treatment as compared to vehicle treatment. The overlapped genes were defined as “rescue DEGs (upregulated)” (left). Ring-heatmap plot showing the relative expression levels of “rescue DEGs (upregulated)” in mouse myocardial infarction model (right). Bottom, Venn diagram showing the upregulated genes upon myocardial infarction and downregulated genes upon uridine treatment as compared to vehicle treatment. The overlapped genes were defined as “rescue DEGs (downregulated)” (left). Ring-heatmap plot showing the relative expression levels of “rescue DEGs (downregulated)” in mouse myocardial infarction model (right). The color key from blue to amaranth indicates log2(fold change) values from low to high. f Top, GO term and pathway enrichment analysis of “rescue DEGs (upregulated)” in mouse myocardial infarction model. Bottom, GO term and pathway enrichment analysis of “rescue DEGs (downregulated)” in mouse myocardial infarction model. The color keys from white to red or blue indicate –log10(P-value) from low to high.
Additionally, the uridine treatment improved the function of the heart underwent myocardial infraction, as evidenced by elevated left ventricular ejection fraction (LVEF) and left ventricular fractional shortening (LVFS) (Fig. 5a, b). Serum lactate dehydrogenase (LDH) and creatine kinase (CK), leakage of which are indicators of acute myocardial infarction, were also lower in uridine-treated mice than control mice (Fig. 5c). Compared to vehicle-treated mice, global gene expression was also reset to be close to the state before injury in uridine-treated mice, with increased expression of genes related to “heart contraction”, and “cardiac muscle tissue development”, and decreased expression of genes associated with “inflammatory response” (Fig. 5d–f). Altogether, uridine promotes the course of tissue regeneration probably by modulating the metabolic process and suppressing inflammation.
In addition to muscular and cardiac injury models, uridine treatment also facilitates the regeneration of the liver after carbon tetrachloride (CCl4) induced injury as evidenced by increased liver-to-body weight ratio and decreased liver fibrosis (Fig. 6a–c). Meanwhile, liver function was restored to a physiological level, such as the total bile acid production (Fig. 6d). In the hair regeneration model, we found that uridine supplementation initiated a new wave of hair growth, as revealed by actively cycling hair follicles with high expression of the proliferation marker Ki67 upon uridine supplementation (Fig. 6e–i). In another tissue injury model, uridine treatment facilitated the regeneration of injured cartilage as assessed by safranin O-fast green staining and further ameliorated functional deterioration, as shown by improved grip strength and athletic ability compared to those of the vehicle-treated group (Fig. 6j–m). Finally, we evaluated the effect of uridine supplementation in physiologically aged mice (22 months old) (Fig. 6n–p) and found improved locomotive activities in the mice with oral administration of uridine for 2 months, as indicated by their enhanced grip strength and exercise endurance (Fig. 6o, p). Overall, by combining systematic metabolomics analysis across multiple models with small-molecule screening for regenerative activity, we identified the endogenous small-molecule metabolite uridine as an effective compound that promotes the repair and regeneration of various tissues and organs, which has the potential to extend the healthspan of aged individuals (Supplementary Fig. S5e).

a Schematic diagram for the experimental design of the mouse liver fibrosis (LF) modeling and vehicle or uridine treatment. b Bar charts of liver weight (left) and liver index (right) of sham mice (n = 10 mice) and liver fibrotic mice treated with vehicle (n = 9 mice) or uridine (n = 10 mice). c Representative images of Masson staining of the liver from sham mice (n = 10 mice) and liver fibrotic mice treated with vehicle (n = 9 mice) or uridine (n = 10 mice). Quantitative data of the relative fibrotic area are shown to the right. Scale bars, 200 μm. d Diagnostic tests for liver functions of sham mice (n = 10 mice) and liver fibrotic mice treated with vehicle (n = 9 mice) or uridine (n = 10 mice). e Schematic diagram for the time course of the mouse hair regeneration experiment. f Hair-growth effect of mice topically treated or subcutaneously injected with vehicle or uridine. Hair-growth rates upon vehicle or uridine treatment were verified by pigmentation scoring. Subcutaneous injection (Vehicle, n = 7 mice; Uridine, n = 8 mice). Topical treatment (Vehicle, n = 6 mice; Uridine, n = 10 mice). g Representative images of H&E staining of the hair follicle of mice subcutaneously injected with vehicle or uridine at day 14 post treatment. Scale bars, 400 μm. h Pie plots showing the hair follicle phase of mice subcutaneously injected with vehicle or uridine at day 14 post treatment. Mean values of hair follicle phases for mice subcutaneously injected with vehicle (n = 7 mice) or uridine (n = 8 mice) are shown. i Ki67 and KRT15 staining of the hair follicle of mice subcutaneously injected with vehicle (n = 7 mice) or uridine (n = 8 mice) at day 14 post treatment. Scale bars, 200 μm. j Schematic diagram for the experimental design of anterior cruciate ligament transection (ACLT) mediated osteoarthritis (OA) modeling and vehicle or uridine treatment. k Representative images of Safranin O/ Fast Green staining of articular cartilage from the joints of sham mice (n = 10 mice) and OA mice treated with vehicle (n = 10 mice) or uridine (n = 10 mice). Quantitative data of cartilage thickness are shown to the right. Scale bars, 100 μm. l Bar chart showing the times of electric shock for sham mice (n = 7 mice) and OA mice treated with vehicle (n = 7 mice) or uridine (n = 7 mice) on the treadmill within 30 min at day 33 post vehicle or uridine treatment. m Bar chart showing the grip strength evaluation of the forelimbs and hind limbs of sham mice (n = 10 mice) and OA mice treated with vehicle (n = 10 mice) or uridine (n = 10 mice) at day 33 post vehicle or uridine treatment. n Schematic diagram of the long-term oral administration experiment. o Bar chart showing the grip strength evaluation of the forelimbs and hind limbs of mice orally administered vehicle (n = 26 mice) or uridine (n = 26 mice) at day 57. p Bar chart showing the times of electric shock for mice orally administered vehicle (n = 26 mice) or uridine (n = 22 mice) on the treadmill within 30 min at day 63 post treatment. Data in b–d, f, i, k–m, o, and p are presented as the means ± SEM. (two-tailed unpaired Student’s t-test).
Discussion
The fundamental questions we try to answer in this study are how high regenerative capacity is fueled by metabolic mechanisms and how we can enhance regeneration through metabolic intervention. By combining metabolomics and transcriptomics approaches to survey phenotypes that are selectively present in actively regenerating tissues and stem cells across species, we shed new light on cross-species and cross-ages metabolic mechanisms associated with regenerative capacity.
In most vertebrates, tissue regeneration is impaired by aging, due to concomitant with cellular senescence, organ degeneration, and other age-associated comorbidities2. The youth factors frequently decreased with age48,49, such as polyamines (spermidine or spermine), have been reported to promote the regeneration of tissues and delay the progression of aging-related disorders50,51,52,53. Metabolic profiling of young tissues and stem cell models with higher regenerative capacity enables us to discover new youth factors and mechanisms associated with regeneration enhancement conserved across species.
The assembly of our metabolomic atlas allowed us to discover metabolic differences between samples with high and low regenerative abilities, but also enabled the identification of dozens of metabolite effectors with the potential to promote tissue regeneration. Here, through the cross-species metabolomics analysis and metabolites screening, we identified endogenous metabolite uridine as a potent regeneration promoting factor. Uridine was identified to be more abundant in tissues or cells with higher regenerative potential. In particular, the concentration of uridine decreased in the plasma from aged individuals, suggesting uridine may navigate a delicate balance between aging and regeneration. In addition, uridine supplementation rejuvenated senescent stem cells, promoted the regeneration and repair of multiple mammalian tissues, and improved the fitness of aged mice. Given the beneficial roles of uridine in promoting tissue repair and improving physiological function, these findings may also have broad relevance for healthy aging treatments. Our study also identified that a single-dose intraperitoneal administration of uridine in mice is sufficient to make the blood uridine concentration reach the concentration required to rejuvenate aged human stem cells in vitro (Fig. 3b and Supplementary Fig. S5d), suggesting a possibility that uridine treatment may result in systemic exposure to uridine to enhance regeneration of different types of tissues in vivo (Supplementary Fig. S5e). Even though the pharmacokinetic study was conducted in rodents, these results also provide useful information for the design of future primate-based preclinical studies and clinical trials on uridine. In a mechanistic view, uridine supplementation reduces inflammation in vitro and in vivo. Supporting our finding, local uridine administration alleviated symptoms of inflammatory bowel disease in mice, concomitant with inhibiting NF-κB signaling54. As a test for safety, our data showed that long-term uridine treatment via intraperitoneal injection (up to 5 months) or oral administration (up to 7 months) was nontumorigenic. In fact, beneficial roles of uridine metabolism and uracil analogs in cancer treatment have also been reported55,56.
In summary, our study reveals previously unknown metabolism-linked regeneration principles across different species, serving as a mineable resource for investigating regenerative pathways and geroprotective metabolic factors with broad translational potential.
Materials and methods
Animal housing and tissue sampling
The use of cynomolgus monkeys and mice in this study was approved by the Ethics Review Committee of the Institute of Zoology, Chinese Academy of Sciences. Monkeys originating from Southeast Asia were housed in cages under a 12-h light-dark cycle at the certified Primate Research Center in Bei**g (** quality more than 20) were further analyzed86. Differentially expressed genes (DEGs) were calculated by R package DESeq2 (version 1.22.2) with a cutoff of Benjamini-Hochberg adjusted P-value < 0.05 and |log2(fold change)| > 0.25 for axolotl tissues, hMSCs, and mice tissues and P-value < 0.05 and |log2(fold change)| > 0.25 for NHP tissues87. The annotation for axolotl genes was conducted following a previous study21. Downregulated genes in injured muscle or heart samples and then restored upon uridine treatment were termed as “rescue DEGs (upregulated)”, and upregulated genes in injured muscle or heart samples and then restored upon uridine treatment were termed as “rescue DEGs (downregulated)”.
Gene ontology (GO), pathway and gene set analysis
GO and pathway enrichment analyses were conducted using Metascape88.
Gene set enrichment analysis was conducted using GSEA (version 4.1.0) with default parameters89.
The differential expression (DE) score for the indicated pathway was calculated as
Mitochondria-localized genes were obtained from MitoCarta database (version 3.0)90.
Regeneration-associated genes were obtained from REGene database91.
Metabolic genes were obtained from Kyoto Encyclopedia of Genes and Genomes (KEGG) database92.
Transcription factor (TF) enrichment analysis
Transcription factor enrichment analysis was performed using R package RcisTarget (version 1.10.0)93. The transcription factor networks were visualized with Cytoscape (version 3.8.0)94.
Metabolic pathway enrichment analysis with transcriptomic and metabolomic data
The integrated pathway-level analysis of transcriptomic and metabolomic data was conducted with “Joint Pathway Analysis” module in MetaboAnalyst (version 4.0)39.
Analysis of snRNA-seq data
snRNA-seq data were processed with Cell Ranger (version 3.1.0). The pre-mRNA Mus musculus (version mm10) reference was built following the Cell Ranger protocol (https://support.10xgenomics.com/single-cell-gene-expression/software/pipelines/latest/advanced/references).
The expression matrix from Cell Ranger output was calculated with Seurat (version 3.1.3)95. Cells with more than 200 genes and with a mitochondrial gene ratio of fewer than 5% were kept. Doublets were identified using DoubletFinder (version 2.0.2)96. Upon normalization and clustering as described below, a cluster with relatively higher mitochondrial gene ratios and lower unique molecular identifier (UMI) counts were also excluded. Finally, 22,713 high-quality cells were retained for downstream analyses.
Dataset of each sample was normalized by “SCTransfrom” function of Seurat. After datasets from different samples were integrated and scaled, the principal component analysis was performed with the “RunPCA” function, and clusters were then identified using the “FindClusters” function. Dimensionality reduction was conducted with the “RunUMAP” function. Differential gene expression analysis was performed using the “FindMarkers” function of Seurat between different groups with the Wilcox test. DEGs were identified as those with adjusted P-value < 0.05 and |log2(fold change)| > 0.25.
Statistical analysis
Data are presented as the means ± SEM. P-value (P) was calculated using GraphPad Prism 9 software by two-tailed Student’s t-test. Statistical methods are indicated in figure legends.
Data availability
The raw sequence data of RNA-seq generated in this study have been deposited in the Genome Sequence Archive in National Genomics Data Center under accession numbers CRA002966, CRA002968, CRA002965, CRA003067, CRA003064, CRA004077, and CRA004294. The differentially present metabolic products and differentially expressed genes identified in our regenerative or uridine-treated systems have been deposited to the Regeneration Roadmap (RR, https://ngdc.cncb.ac.cn/aging/index) or Aging Atlas (AA, https://ngdc.cncb.ac.cn/aging/index) databases97,98.
References
Jopling, C., Boue, S. & Izpisua Belmonte, J. C. Dedifferentiation, transdifferentiation and reprogramming: three routes to regeneration. Nat. Rev. Mol. Cell Biol. 12, 79–89 (2011).
Yun, M. H. Changes in regenerative capacity through lifespan. Int J. Mol. Sci. 16, 25392–25432 (2015).
McCusker, C., Bryant, S. V. & Gardiner, D. M. The axolotl limb blastema: cellular and molecular mechanisms driving blastema formation and limb. Regeneration tetrapods. Regeneration (Oxf.) 2, 54–71 (2015).
Phan, A. Q. et al. Positional information in axolotl and mouse limb extracellular matrix is mediated via heparan sulfate and fibroblast growth factor during limb regeneration in the axolotl (Ambystoma mexicanum). Regeneration (Oxf.) 2, 182–201 (2015).
Endo, T., Bryant, S. V. & Gardiner, D. M. A stepwise model system for limb regeneration. Dev. Biol. 270, 135–145 (2004).
Li, C. Deer antler regeneration: a stem cell-based epimorphic process. Birth Defects Res C. Embryo Today 96, 51–62 (2012).
Wang, D. et al. Deer antler stem cells are a novel type of cells that sustain full regeneration of a mammalian organ-deer antler. Cell Death Dis. 10, 443 (2019).
Li, C., Zhao, H., Liu, Z. & McMahon, C. Deer antler – A novel model for studying organ regeneration in mammals. Int. J. Biochem. Cell Biol. 56, 111–122 (2014).
Neves, J., Sousa-Victor, P. & Jasper, H. Rejuvenating strategies for stem cell-based therapies in aging. Cell Stem Cell 20, 161–175 (2017).
West, M. D. et al. Toward a unified theory of aging and regeneration. Regen. Med. 14, 867–886 (2019).
Di Micco, R., Krizhanovsky, V., Baker, D. & d’Adda di Fagagna, F. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat. Rev. Mol. Cell Biol. 22, 75–95 (2021).
Yao, M. et al. PCGF5 is required for neural differentiation of embryonic stem cells. Nat. Commun. 9, 1463 (2018).
Gerber, T. et al. Single-cell analysis uncovers convergence of cell identities during axolotl limb regeneration. Science 362, eaaq0681 (2018).
Dong, Z., Coates, D., Liu, Q., Sun, H. & Li, C. Quantitative proteomic analysis of deer antler stem cells as a model of mammalian organ regeneration. J. Proteom. 195, 98–113 (2019).
Wang, Y. et al. Genetic basis of ruminant headgear and rapid antler regeneration. Science 364, eaav6335 (2019).
Lei, J. et al. Exosomes from antler stem cells alleviate mesenchymal stem cell senescence and osteoarthritis. Protein Cell https://doi.org/10.1007/s13238-021-00860-9 (2021).
Ichu, T.-A., Han, J., Borchers, C. H., Lesperance, M. & Helbing, C. C. Metabolomic insights into system-wide coordination of vertebrate metamorphosis. BMC Dev. Biol. 14, 5 (2014).
Shen, B. et al. Proteomic and metabolomic characterization of COVID-19 patient sera. Cell 182, 59–72 (2020).
Tanaka, E. M. Regenerating tissues. Science 360, 374–375 (2018).
Bryant, S. V. & Gardiner, D. M. Regeneration: sooner rather than later. Int J. Dev. Biol. 62, 363–368 (2018).
Li, H. et al. Dynamic cell transition and immune response landscapes of axolotl limb regeneration revealed by single-cell analysis. Protein Cell 12, 57–66 (2021).
Zhang, W. et al. Aging stem cells. A Werner syndrome stem cell model unveils heterochromatin alterations as a driver of human aging. Science 348, 1160–1163 (2015).
Wu, Z. et al. Differential stem cell aging kinetics in Hutchinson-Gilford progeria syndrome and Werner syndrome. Protein Cell 9, 333–350 (2018).
Geng, L. et al. Chemical screen identifies a geroprotective role of quercetin in premature aging. Protein Cell 10, 417–435 (2019).
Wang, W. et al. A genome-wide CRISPR-based screen identifies KAT7 as a driver of cellular senescence. Sci. Transl. Med. 13, eabd2655 (2021).
Liu, X. et al. Resurrection of human endogenous retroviruses during aging reinforces senescence. bioRxiv https://doi.org/10.1101/2021.02.22.432260 (2021).
Shan, H. et al. Large-scale chemical screen identifies Gallic acid as a geroprotector for human stem cells. Protein Cell https://doi.org/10.1007/s13238-021-00872-5 (2021).
Aoyama, T. et al. Cloning of human very-long-chain acyl-coenzyme A dehydrogenase and molecular characterization of its deficiency in two patients. Am. J. Hum. Genet. 57, 273–283 (1995).
McAndrew, R. P. et al. Structural basis for substrate fatty acyl chain specificity: crystal structure of human very-long-chain acyl-CoA dehydrogenase. J. Biol. Chem. 283, 9435–9443 (2008).
Zhu, J. & Thompson, C. B. Metabolic regulation of cell growth and proliferation. Nat. Rev. Mol. Cell Biol. 20, 436–450 (2019).
Solhi, R., Lotfinia, M., Gramignoli, R., Najimi, M. & Vosough, M. Metabolic hallmarks of liver regeneration. Trends Endocrinol. Metab. 32, 731–745 (2021).
Khalafalla, F. G. et al. P2Y2 nucleotide receptor prompts human cardiac progenitor cell activation by modulating hippo signaling. Circ. Res. 121, 1224–1236 (2017).
Wu, G., Fang, Y.-Z., Yang, S., Lupton, J. R. & Turner, N. D. Glutathione metabolism and its implications for health. J. Nutr. 134, 489–492 (2004).
Vats, P., Singh, V. K., Singh, S. N. & Singh, S. B. Glutathione metabolism under high-altitude stress and effect of antioxidant supplementation. Aviat. Space Environ. Med. 79, 1106–1111 (2008).
Colemonts-Vroninks, H. et al. Oxidative Stress, Glutathione Metabolism, and Liver Regeneration Pathways Are Activated in Hereditary Tyrosinemia Type 1 Mice upon Short-Term Nitisinone Discontinuation. Genes 12, 3 (2021).
Doi, J. et al. Bolus administration of polyamines boosts effects on hepatic ischemia-reperfusion injury and regeneration in rats. Eur. Surg. Res. 60, 63–73 (2019).
Sanchez-Sevilla, L., Mendieta-Condado, E. & Hernandez-Munoz, R. Putrescine treatment reverses alpha-tocopherol-induced desynchronization of polyamine and retinoid metabolism during rat liver regeneration. J. Transl. Med. 14, 307 (2016).
Madeo, F., Eisenberg, T., Pietrocola, F. & Kroemer, G. Spermidine in health and disease. Science 359, eaan2788 (2018).
Chong, J. et al. MetaboAnalyst 4.0: towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 46, W486–W494 (2018).
Amber-Vitos, O., Chaturvedi, N., Nachliel, E., Gutman, M. & Tsfadia, Y. The effect of regulating molecules on the structure of the PPAR-RXR complex. Biochim Biophys. Acta. 1861, 1852–1863 (2016).
Plutzky, J. The PPAR-RXR transcriptional complex in the vasculature: energy in the balance. Circ. Res. 108, 1002–1016 (2011).
van Neerven, S., Kampmann, E. & Mey, J. RAR/RXR and PPAR/RXR signaling in neurological and psychiatric diseases. Prog. Neurobiol. 85, 433–451 (2008).
Yan, P. et al. FOXO3-engineered human ESC-derived vascular cells promote vascular protection and regeneration. Cell Stem Cell 24, 447–461.e8 (2019).
Zhang, W. et al. A single-cell transcriptomic landscape of primate arterial aging. Nat. Commun. 11, 2202 (2020).
Miret-Casals, L. et al. Identification of new activators of mitochondrial fusion reveals a link between mitochondrial morphology and pyrimidine metabolism. Cell Chem. Biol. 25, 268–278.e4 (2018).
Eming, S. A., Wynn, T. A. & Martin, P. Inflammation and metabolism in tissue repair and regeneration. Science 356, 1026–1030 (2017).
Biferali, B., Proietti, D., Mozzetta, C. & Madaro, L. Fibro-adipogenic progenitors cross-talk in skeletal muscle: the social network. Front. Physiol. 10, 1074 (2019).
Nishimura, K., Shiina, R., Kashiwagi, K. & Igarashi, K. Decrease in polyamines with aging and their ingestion from food and drink. J. Biochem. 139, 81–90 (2006).
Minois, N., Carmona-Gutierrez, D. & Madeo, F. Polyamines in aging and disease. Aging 3, 716–732 (2011).
Noro, T. et al. Spermidine promotes retinal ganglion cell survival and optic nerve regeneration in adult mice following optic nerve injury. Cell death Dis. 6, e1720–e1720 (2015).
Wang, J. et al. Spermidine alleviates cardiac aging by improving mitochondrial biogenesis and function. Aging (Albany NY) 12, 650–671 (2020).
Xu, T.-T. et al. Spermidine and spermine delay brain aging by inducing autophagy in SAMP8 mice. Aging 12, 6401–6414 (2020).
Minguzzi, M. et al. Polyamine supplementation reduces DNA damage in adipose stem cells cultured in 3-D. Sci. Rep. 9, 14269 (2019).
Jeengar, M. K., Thummuri, D., Magnusson, M., Naidu, V. G. M. & Uppugunduri, S. Uridine ameliorates dextran sulfate sodium (DSS)-induced colitis in mice. Sci. Rep. 7, 3924 (2017).
Field, M. S., Lan, X., Stover, D. M. & Stover, P. J. Dietary uridine decreases tumorigenesis in the Apc(Min/+) model of intestinal cancer. Curr. Dev. Nutr. 2, nzy013 (2018).
Ma, W. W. et al. Emergency use of uridine triacetate for the prevention and treatment of life-threatening 5-fluorouracil and capecitabine toxicity. Cancer 123, 345–356 (2017).
Zhang, H. et al. Single-nucleus transcriptomic landscape of primate hippocampal aging. Protein Cell https://doi.org/10.1007/s13238-021-00852-9 (2021).
Wang, S. et al. Deciphering primate retinal aging at single-cell resolution. Protein Cell https://doi.org/10.1007/s13238-020-00791-x (2020).
Li, J. et al. A single-cell transcriptomic atlas of primate pancreatic islet aging. Natl. Sci. Rev. 8, https://doi.org/10.1093/nsr/nwaa127 (2021).
Ling, C. et al. Modeling CADASIL vascular pathologies with patient-derived induced pluripotent stem cells. Protein Cell 10, 249–271 (2019).
Wang, S. et al. Rescue of premature aging defects in Cockayne syndrome stem cells by CRISPR/Cas9-mediated gene correction. Protein Cell 11, 1–22 (2020).
Zhang, X. et al. Telomere-dependent and telomere-independent roles of RAP1 in regulating human stem cell homeostasis. Protein Cell 10, 649–667 (2019).
Wang, S. et al. ATF6 safeguards organelle homeostasis and cellular aging in human mesenchymal stem cells. Cell Discov. 4, 2 (2018).
Bi, S. et al. SIRT7 antagonizes human stem cell aging as a heterochromatin stabilizer. Protein Cell 11, 483–504 (2020).
Liang, C. et al. Stabilization of heterochromatin by CLOCK promotes stem cell rejuvenation and cartilage regeneration. Cell Res. 31, 187–205 (2021).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Cheng, F. et al. DJ-1 is dispensable for human stem cell homeostasis. Protein Cell 10, 846–853 (2019).
Pan, H. et al. SIRT6 safeguards human mesenchymal stem cells from oxidative stress by coactivating NRF2. Cell Res. 26, 190–205 (2016).
Alves, A. N. et al. Comparative effects of low-level laser therapy pre- and post-injury on mRNA expression of MyoD, myogenin, and IL-6 during the skeletal muscle repair. Lasers Med Sci. 31, 679–685 (2016).
Gao, E. et al. A novel and efficient model of coronary artery ligation and myocardial infarction in the mouse. Circ. Res. 107, 1445–1453 (2010).
Lei, J. et al. FOXO3-engineered human mesenchymal progenitor cells efficiently promote cardiac repair after myocardial infarction. Protein Cell 12, 145–151 (2021).
Chai, M. et al. Stimulation of hair growth by small molecules that activate autophagy. Cell Rep. 27, 3413–3421.e3 (2019).
Oh, J. W. et al. A guide to studying human hair follicle cycling in vivo. J. Invest Dermatol 136, 34–44 (2016).
Bataller, R. & Brenner, D. A. Liver fibrosis. J. Clin. Invest 115, 209–218 (2005).
Sakaida, I. et al. Transplantation of bone marrow cells reduces CCl4-induced liver fibrosis in mice. Hepatology 40, 1304–1311 (2004).
Deng, L. et al. Stabilizing heterochromatin by DGCR8 alleviates senescence and osteoarthritis. Nat. Commun. 10, 3329 (2019).
Fu, L. et al. Up-regulation of FOXD1 by YAP alleviates senescence and osteoarthritis. PLoS Biol. 17, e3000201 (2019).
Ren, X. et al. Maintenance of Nucleolar Homeostasis by CBX4 Alleviates Senescence and Osteoarthritis. Cell Rep. 26, 3643–3656.e7 (2019).
Gorkin, D. U. et al. An atlas of dynamic chromatin landscapes in mouse fetal development. Nature 583, 744–751 (2020).
Krishnaswami, S. R. et al. Using single nuclei for RNA-seq to capture the transcriptome of postmortem neurons. Nat. Protoc. 11, 499–524 (2016).
Ma, S. et al. Caloric Restriction Reprograms the Single-Cell Transcriptional Landscape of Rattus Norvegicus Aging. Cell 180, 984–1001.e2 (2020).
Ma, S. et al. Single-cell transcriptomic atlas of primate cardiopulmonary aging. Cell Res. 31, 415–432 (2021).
Hakimi, A. A. et al. An inte grated metabolic atlas of clear cell renal cell carcinoma. Cancer Cell 29, 104–116 (2016).
Yan, P. et al. Genome-wide R-loop landscapes during cell differentiation and reprogramming. Cell Rep. 32, 107870 (2020).
Kim, D., Langmead, B. & Salzberg, S. L. HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12, 357 (2015).
Anders, S., Pyl, P. T. & Huber, W. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Zhou, Y. et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 10, 1523 (2019).
Subramanian, A., Kuehn, H., Gould, J., Tamayo, P. & Mesirov, J. P. GSEA-P: a desktop application for Gene Set Enrichment Analysis. Bioinformatics 23, 3251–3253 (2007).
Rath, S. et al. MitoCarta3.0: an updated mitochondrial proteome now with sub-organelle localization and pathway annotations. Nucleic Acids Res. 49, D1541–D1547 (2021).
Zhao, M., Rotgans, B., Wang, T. & Cummins, S. F. REGene: a literature-based knowledgebase of animal regeneration that bridge tissue regeneration and cancer. Sci. Rep. 6, 23167 (2016).
Kanehisa, M., Sato, Y. & Kawashima, M. KEGG map** tools for uncovering hidden features in biological data. Protein Sci https://doi.org/10.1002/pro.4172 (2021).
Aibar, S. et al. SCENIC: single-cell regulatory network inference and clustering. Nat. Methods 14, 1083–1086 (2017).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Butler, A., Hoffman, P., Smibert, P., Papalexi, E. & Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420 (2018).
McGinnis, C. S., Murrow, L. M. & Gartner, Z. J. DoubletFinder: doublet detection in single-cell RNA sequencing data using artificial nearest neighbors. Cell Syst. 8, 329–337.e4 (2019).
Aging Atlas, C. Aging Atlas: a multi-omics database for aging biology. Nucleic Acids Res. 49, D825–D830 (2021).
Kang, W. et al. Regeneration Roadmap: database resources for regenerative biology. Nucleic Acids Res. https://doi.org/10.1093/nar/gkab870 (2021).
Acknowledgements
The authors thank Qianzhao Ji, Wang Kang, Yandong Zheng and Shuai Ma for their assistance in the bioinformatic analysis. We would like to thank Yifang He, Li** Deng, Ruotong Ren, Yanling Fan, Ji** Yang, Wenlei Bao, Zhiheng He and **ng Zhang for participating in the organ and tissue harvesting from anesthetized monkeys. We thank Yuhang Zhang, Yanxia Ye for their help in cell culture. We are also grateful to Lei Bai, **g Lu, Ying Yang, Ruijun Bai, Lei Zhang, Luyang Tian, and Shikun Ma for administrative assistance, and Junying Jia from the Institute of Biophysics, Chinese Academy of Sciences for his help with the fluorescence-activated cell sorting (FACS) and Junfeng Hao from the Institute of Biophysics for her help in image scanning and analysis of H&E staining. We thank Yan Teng and Chunliu Liu from Center for Biological Imaging, Institute of Biophysics, Chinese Academy of Sciences for their help of taking confocal images. We thank **ang Shi, Lei Zhou, **nyi Wu, Zhuanzhuan **ng, **mei Zhang, Sai Yang, Yongxia Liu, Jiajia Hou, Shuguang Duo, Qiong Jia, Yue **e, Xuancheng Lu, **aomeng Shi, Shuai Pang, **aoyu Wang, Yukun Wang and Yakun Chen for management of laboratory animals. This work was supported by the National Key Research and Development Program of China (2020YFA0804000), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA16010000), the National Key R&D Program of China (2018YFC2000100, 2017YFA0103300, 2017YFA0102800, 2018YFA0107200, 2020YFA0113400, 2020YFA0112200, 2019YFA0110100, 2020YFA0803401, 2019YFA0802202, 2018YFC2000400, 2021YFF1201000), the National Natural Science Foundation of China (82125011, 81921006, 81625009, 91749202, 81861168034, 91949209, 92049304, 81822018, 92049116, 81870228, 81922027, 31970597, 82071588, 82122024, 31801010, 82001477, 81801370, 32100937, 81901433, 81801399, 92149301, 92168201), the Program of the Bei**g Municipal Science and Technology Commission (Z191100001519005), Bei**g Natural Science Foundation (Z190019, JQ20031), the Key Research Program of the Chinese Academy of Sciences (KFZD-SW-221), the 14th Five-year Network Security and Informatization Plan of Chinese Academy of Sciences (WX145XQ07-18), K. C. Wong Education Foundation (GJTD-2019-06, GJTD-2019-08), Bei**g Hospitals Authority Youth Programme (QML20200802),Youth Innovation Promotion Association of CAS (E1CAZW0401, 2020085, 2021078), the Informatization Plan of Chinese Academy of Sciences (CAS-WX2021SF-0301), the CAS Project for Young Scientists in Basic Research (YSBR-012), the Non-profit Central Research Institute Fund of Chinese Academy of Medical Sciences (2020-JKCS-011), Young Elite Scientists Sponsorship Program by CAST(YESS20200012), CAS Special Research Assistant (SRA) Program, Priority Union Foundation of Yunnan Provincial Science and Technology Department (202001AY070001-011), the State Key Laboratory of Stem Cell and Reproductive Biology, and the State Key Laboratory of Membrane Biology and the Milky Way Research Foundation (MWRF).
Author information
Authors and Affiliations
Contributions
G.-H.L., J.Q., and W.Z., conceptualized this project and supervised the overall experiments. Z.L. performed bioinformatics analysis of the metabolomics data and bulk RNA-seq data. W.L., J.L., Q.Z., Y.C., and Q.C. performed mouse experiments, immunostaining and tissue section analyses. L.G. performed cell-based metabolite screening. P.Y. and C.L. performed in vitro uridine treatment assays and phenotype analysis. S.W. and Y.C. performed single-nucleus isolation of mice muscles. J.L. performed bioinformatics analysis of snRNA-seq data. Q.W. and C.L. performed amputation and tissue collection of Mexican axolotl. L.S. provided human plasma. Q.W. drew the schematic illustration. G.-H.L., J.Q., W.Z., Z.L., W.L., L.G., L.S., Y.Y., P.Y., C.L., J.R., M.S., K.Y., Z.W., J.L., S.W., C.L., J.-D.J.H., R.H., N.S.C., and J.C.I.B. performed manuscript writing, review, and editing.
Corresponding authors
Ethics declarations
Conflict of interest
The author declares no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, Z., Li, W., Geng, L. et al. Cross-species metabolomic analysis identifies uridine as a potent regeneration promoting factor. Cell Discov 8, 6 (2022). https://doi.org/10.1038/s41421-021-00361-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41421-021-00361-3
- Springer Nature Singapore Pte Ltd.
We’re sorry, something doesn't seem to be working properly.
Please try refreshing the page. If that doesn't work, please contact support so we can address the problem.