Abstract
Pancreatic cancer (PC) is the most hypoxic cancer type among solid tumors. The dynamic changes of RNA N6-methyl-adenosine (m6A) contribute to tumor cells adaption to hypoxic microenvironmental. However, the regulatory mechanisms of hypoxia response in PC remains elusive. Here, we reported that the m6A demethylase ALKBH5 mediated a decrease of total mRNA m6A modification during hypoxia. Subsequently, methylated RNA immunoprecipitation sequencing (MeRIP-seq) combined with RNA sequencing (RNA-seq) revealed transcriptome-wide gene expression alteration and identified histone deacetylase type 4 (HDAC4) as a key target gene of m6A modification under hypoxic conditionds. Mechanistically, m6A methylation recognized by m6A reader-YTHDF2 enhanced the stability of HDAC4, and then promoted glycolytic metabolism and migration of PC cells. Our assays also demonstrated that hypoxia-induced HDAC4 enhanced HIF1a protein stability, and overexpressed HIF1a promoted transcription of ALKBH5 in hypoxic pancreatic cancer cells. Together, these results found a ALKBH5/HDAC4/HIF1α positive feedback loop for cellular response to hypoxia in pancreatic cancer. Our studies uncover the crosstalk between histone acetylation and RNA methylation modification on layer of epigenetic regulation.
Similar content being viewed by others


Introduction
Pancreatic cancer (PC) is the malignant tumor with the highest mortality rate among gastrointestinal tumors. It is characterized by insignificant initial symptoms, rapid deterioration, low surgical resection rate, and poor overall prognosis [31]. In this study, we found the methylation level of HDAC4 mRNA was decreased under hypoxia and the expression of HDAC4 was increased. That is, m6A modification negatively regulates HDAC4 expression. According to the different functions of “reader” proteins, we speculate that YTHDF2 may be involved in the regulation of HDAC4 expression by regulating its mRNA stability. To substantiate this hypothesis, we firstly measured the expression level of YTHDC2 and YTHDF2, and the western blot assay showed, those “readers” were not affected under hypoxic conditions (Fig. 4E). Furthermore, we established YTHDF2-silenced stable cell lines (Supplementary Fig. 4A, B). Transcriptional expression level of HDAC4 were detected using a qRT-PCR assay, we found that HDAC4 mRNA expression levels were significantly increased when YTHDF2 was impaired compared with those in control cells (Fig. 4F). Thereafter, we performed RIP assays with anti-YTHDF2 antibody, as detected, HDAC4 mRNA was significantly enriched in the YTHDF2 antibody pull down sample compared with the IgG control (Fig. 4G). Together, our findings indicated that HDAC4 was governed by m6A modification depending on ALKBH5 and recognized by YTHDF2, which receded its stability under hypoxic conditions.
HDAC4 is identified as an oncogenic driver in PC
It has been reported that histone deacetylases (HDACs) promote tumour cell migration and the inhibition of HDACs may be a promising antitumour therapy for pancreatic cancer [32, 33]. Emerging evidence also indicates that HDAC4 mediates smoking-induced pancreatic cancer metastasis [34]. In clinical studies, elevated HDAC4 expression was significantly associated with the absence of organ metastases and tumor proliferative capacity in pancreatic adenocarcinoma [35]. These studies suggest that HDAC4 may be implicated in pancreatic malignant disease progression. In this study, we decided to systematically investigate the function of HDAC4 in pancreatic cancer under hypoxia. First, we established HDAC4-overexpression PANC-1 and MIA PaCa-2cell lines, and cultured them in 20 and 1% oxygen for 48 h (Supplementary Fig. 5A, B). We found that HDAC4 overexpression resulted in the significantly increased proliferation rate of PC cells under hypoxia compared to normoxia (Fig. 5A). The migratory assays indicated that HDAC4 overexpression significantly facilitated migratory ability of PC cells in normoxia and hypoxia (Fig. 5B, Supplementary Fig. 5C, D). Besides, HDAC4 overexpression amplified the hypoxia-induced key glycolysis regulatory genes, such as LDHA, HK2, GLUT1 and GLUT3 (Fig. 5C). Moreover, HDAC4 overexpression also increased production of lactate under normoxic and hypoxic conditions (Fig. 5D, E and Supplementary Fig. 5E). The extracellular acidification rate kinetic profiles also demonstrated the increase of glycolytic activity in HDAC4-overexpression PC cells (Fig. 5F, G and Supplementary Fig. 5F). Those data strongly suggest that HDAC4 could maintain hypoxia-induced glycolysis and metastasis in PC cells.

A PC cells with NC and HDAC4 overexpression were incubated under normoxia or hypoxia. CCK8 assays were used to measure cell growth, and the relative proliferation rates were calculated. B Transwell assays were used to investigate the changes in the migratory capability of PC cells after viral transfection with HDAC4, and cells were exposed to 20% O2 or 1% O2 for 48 h. C PANC-1 and MIA PaCa-2 cells with NC and HDAC4 overexpression were cultured under normoxia or hypoxia for 48 h. And the mRNA expression of glycolysis driver genes (LDHA, HK2, GLUT1 and GLUTG3) was detected by qRT-PCR assay. Error bars are mean ± SEM. (n = 3). D, E Lactate production was measured in HDAC4-overexpressing PANC-1 or MIA PaCa-2 cells incubated under normoxia or hypoxia. F, G The extracellular acidification rate (ECAR) assays were performed to measure the glycolytic metabolism level in HDAC4-overexpressing cells incubated under normoxia or hypoxia. The quantification of glycolysis and glycolytic capacity was summarized from raw data. The metabolic inhibitors were injected sequentially at different time points as indicated. n = 3-4 for each treatment group. *P < 0.05, **P < 0.01, ***P < 0.001 and ns not significant.
Hypoxia-induced glycolysis and metastasis were reversed by silenced HDAC4 in a m6A dependent manner
To confirm that the observed phenotypes were mediated by the dysregulation of ALKBH5- HDAC4 axis, we generated PC cells interfered with HDAC4 (Supplementary Fig. 6A, B). As expected, knockdown of HDAC4 significantly abolished the increased mobility ability induced by hypoxia in ALKBH5-overexpressing cells (Fig. 6A and Supplementary Fig. 6C). The Seahorse Extracellular Flux analysis showed hypoxia enhanced glycolytic capacity of ALKBH5-overexpressing cells, which could be reverted by HDAC4 silencing (Fig. 6B, C). HDAC4 silencing also abolished hypoxia-induced mRNA upregulation of key glycolysis regulatory gene in ALKBH5-overexpressing cells (Fig. 6D, E). And analogous results could be verified in ALKBH5-overexpressing cells, which were treated with tasquinimod (an inhibitor targeting HDAC4) under hypoxia (Fig. 6F–H and Supplementary Fig. 6D). These several functional rescue assays suggested that effects of hypoxia were reversed by silenced HDAC4.

A ALKBH5-overexpressing PC cells transfected with the shNC or sh-HDAC4 were incubated under hypoxic conditions. Transwell assay was used to investigate the cell migration abilities. B, C ALKBH5-overexpressing PC cells transfected with the shNC or sh-HDAC4 were cultured under hypoxic conditions. The ECAR was measured to determine the glycolytic metabolism of treatment cells. Glycolytic variations (right) of glycolysis and glycolytic capacity were summarized from raw data. The metabolic inhibitors were injected sequentially at different time points as indicated. n = 3-4 for each treatment group. D, E ALKBH5-overexpressing PC cells transfected with the shNC or sh-HDAC4 were incubated under hypoxic conditions. The expression of a panel of glucose metabolism-related genes was detected by qRT-PCR. Error bars are mean ± SEM. (n = 3). F ALKBH5-overexpressing PANC-1 and MIA PaCa-2 cells treated with Tasquinimod under hypoxic conditions. The cell migration ability was assessed by Transwell assay. G ALKBH5-overexpressing PANC-1 and MIA PaCa-2 cells treated with Tasquinimod under hypoxic conditions. The glycolytic metabolism of treatment cells was measured by Seahorse XF24 system. Glycolytic variations (right) of glycolysis and glycolytic capacity were summarized from raw data. The metabolic inhibitors were injected sequentially at different time points as indicated. n = 3-4 for each treatment group. H The expression of a panel of glucose metabolism-related genes in PANC-1 and MIA PaCa-2 cells was detected by qRT-PCR. Error bars are mean ± SEM. (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001 and ns not significant.
HDAC4 enhanced the stability of HIF1α in hypoxic pancreatic cancer cells
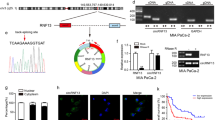
HDAC4 possesses histone deacetylase activity and represses transcription when tethered to a promoter. Recent studies show that HDAC4-HIF1α axis as is an important signal pathway regulating glycolysis, apoptosis and autophagy in ovarian tumor cells during hypoxic adaptation [9, 36]. HDAC4 inhibits HIF1a ubiquitination and enhances its stability through deacetylation. Congruently, in this study, we aimed to demonstrate whether HDAC4 could enhance the stability of HIF1a in hypoxic pancreatic cancer cells. To confirm the regulatory role of HDAC4 on HIF1α, we treated HDAC4-overexpression PANC-1 and MIA PaCa-2 cell lines with Cobalt (II) chloride (CoCl2) to mimic hypoxia condition. Overexpression of HDAC4 significantly delayed HIF1α protein degradation (Fig. 7A). Conversely, knockdown of endogenous HDAC4 facilitated HIF1α protein degradation in hypoxic pancreatic cancer cells (Fig. 7B). We then generated a HIF1α mutant in which the lysine sites affected by HDAC4. The proteasome-specific inhibitor MG132 (10 µM) rescued the HIF1α protein from degradation in HDAC4-knockdown cells. Silenced HDAC4 caused a greater increase in acetylation of the wildtype HIF1α (HIF1α-wt) than in that of the mutant HIF1α (HIF1α-mut) (Fig. 7C). These results suggest that HDAC4 could enhance HIF1α protein stability.

A, B PANC-1 and MIA PaCa-2 cells with HDAC4 overexpression or HDAC4 knockdown were treated with 200 µM CoCl2. Then, the cells were treated with 10 µg/mL cycloheximide (CHX) for the indicated time course (0, 1, 2, 3 h). The protein levels of HIF1α and HDAC4 were analyzed by western blot. C PC cells with or without silencing of HDAC4 expression were transfected with a wild-type Flag-HIF-1α (WT) or mutant Flag-HIF-1α (mut) plasmids for 24 h and exposed to hypoxia for 48 h. Then, the cells were treated with 5 μM MG132 for another 6 h. Cell lysates were immunoprecipitated with an anti-Flag antibody, and then immunoblotted for Flag and the acetylation form of HIF1α. D, E PC cells with HIF1α knockdown (siHIF1α) or control (NC) were treated with or without 200 µM CoCl2, and the expression of HDAC4 and HIF1α was detected using qRT-PCR and western blot. F Predicted HRE sequences in ALKBH5 promoter by JASPAR and the diagram for constructing for constructing of luciferase reporters. The wild-type or HRE mutant sequence of ALKBH5 promoter was inserted into a pcDNA3.1 vector. G Dual luciferase reporter assays showing the effect of HIF1α on ALKBH5 mRNA reporters with either wild-type or mutated HRE sequence under hypoxia. *P < 0.05, **P < 0.01, ***P < 0.001 and ns not significant.
ALKBH5 was transcriptionally induced by HIF1α under hypoxic conditions
It has been reported that ALKBH5 is a nuclear 2-oxoglutarate dependent oxygenase and is a direct target of HIF1α [23, 31], whereas another report showed hypoxia favored m6A labelling in HCC cells [39]. The role of m6A in transcriptome reprogramming during hypoxic response in PC cells are still unclear. In this study, we systematic analyzed the alterations of m6A and its target genes in hypoxic pancreatic cancer cells by the MeRIP-seq. We found that hypoxia reduced m6A modification level of total mRNAs in a ALKBH5 dependent manner. Unexpectedly, our data suggested that ALKBH5 may act as an oncogenic driver to promote cell metastasis in hypoxia instead of serving as a tumor suppressor in normoxia. Nevertheless, the mechanism by which ALKBH5 acts inversely in normoxia and hypoxia is still unclear.
Glycolysis plays a critical role in the regulation of the tumour microenvironment, affecting biological processes such as inflammatory factor secretion, immune evasion and tumor angiogenesis [40,41,42]. Glycolysis is one of the primary metabolic signatures in cancers and supports the energetic requirements of sustained proliferation and metastasis [43]. Apart from DNA and histone modifications, RNA m6A modification has been recently proposed to be another important layer of epigenetic regulation in energy metabolism regulation [44, 31, 47]. HIFs are composed of the stable HIF1β subunit and the oxygen-sensitive HIF1/2α subunit [48]. HIF1α is a crucial regulator of oxygen homeostasis and is degraded by the ubiquitin–proteasome pathway [49, 50]. Under hypoxic conditions, HIF1α is stabilized and binds with HIF1β to promote the transcription of downstream gene [51, 52]. Ling et al. found that the stability and transcriptional activity of HIF1α were enhanced by USP22 depended its deubiquitination activity [50]. Consistent with a previous report [9], we detected HDAC4-mediated deacetylation enhanced HIF1α stability under hypoxia. Recently, ALKBH5 was shown to be directly targeted by HIF1α and regulated by HIF2α in breast cancer cells [38]. Our study also confirmed that overexpressing HIF1α promoted ALKBH5 transcription in pancreatic cancer cells. Therefore, we demonstrated a positive feedback loop between ALKBH5, HDAC4 and HIF1α, which contributes to the maintenance of PC glycolysis under hypoxic microenvironment (Fig. 9).

The expression of ALKBH5 was increased under hypoxic conditions which led to increased stability of HDAC4 mRNA via m6A modification and in a YTHDF2 dependent manner. Increased HDAC4 enhanced HIF1a protein stability. And then overexpressed HIF1a promoted transcription of ALKBH5, LDHA, HK2, GLUT1 and GLUT3. Therefore, ALKBH5/HDAC4/HIF1α form a positive feedback loop, which then induces a more glycolytic metabolism and migration of PC cells under hypoxia.
Altogether, our study revealed the mechanism of m6A modification and the ALKBH5/HDAC4/HIF1α positive feedback loop in promoting glycolysis of PC. Based on our findings, the hypoxia-directed treatment maybe a novel therapeutic option for PC patients.
Materials and methods
RNA m6A dot blot assay
Poly(A)+ mRNA was enriched using GenElute messenger RNA (mRNA) Miniprep Kit (Sigma‐Aldrich). The concentration and purity of RNA were measured by an Agilent Bioanalyzer 2100. Poly(A)+ mRNA samples were denatured at 70 °C for 5 min and then diluted equally to 400 ng, 200 ng and 100 ng in equal volumes. After denaturation, equal volume of diluted mRNA was added into a nylon membrane (GE Healthcare). The membranes were cross-linked at 245 nm UV for three times under auto-cross-linking mode (UVP analytik jena, CL-1000M). Then, the membranes were blocked with 5% BSA for 2 h at room temperature and incubated with anti‐m6A antibody (Synaptic System, 202003) overnight at 4 °C. Membranes were washed with PBST for three times. Horseradish peroxidase-conjugated anti-rabbit immunoglobulin G (CST, 7074P2) were diluted 1:5000 and incubated with the membranes for 1 h at room temperature. Membranes were washed with PBST and the signals were detected by standard analysis of HRPO-induced chemiluminescence (ECL, Millipore). The same gradient-diluted Poly(A)+ mRNA was also added into the nylon membrane, stained with methylene blue (Solarbio, G1300) for 2 h, and washed with ribonuclease-free water.
m6A sequencing and m6A-RNA immunoprecipitation assay
MeRIP sequencing was performed by Jiayin Biomedical Technology company (shanghai, China). In brief, total RNAs were isolated from PANC-1 cells exposed to hypoxia or normoxia, and then chemically fragmented into 100 ~ 200 nt. The fragmented RNA was incubated with m6A antibody (milipore) for immunoprecipitation. The enrichment of m6A containing mRNA was sent for high-throughput NGS and validated by quantitative RT–PCR. For NGS, purified RNA fragments from m6A-MeRIP were used for library construction (removing rRNA). Sequencing reads were aligned to the human genome hg38_gencode with STAR program. The m6A modification peaks were identified using software package of MetPeak. Motifs enriched with m6A peaks were identified by MEME. The R software package Guitar was used to calculate the frequency of each sample’s peak at each site in the mRNA transcript region and draw the frequency distribution map. The distribution of peaks in the functional area covered is drawn by R software package ChiIPseeker. Finally, GO and Pathway enrichment analysis of m6A peaks associated genes were carried out.
Pimonidazole (PIMO) staining assay
2 mg pimonidazole (Hypoxyprobe) was injected intravenously into each tumor-bearing mouse and was left to circulate for 1 h before tumour resection. The obtained tumor tissue was sectioned by LEICA CM1950 (Leica). Staining of tumour sections were performed with a Hypoxyprobe RedAPC Kit (HP8-100) according to manufacturer’s instructions. Specifically, tumour sections were fixed in 4% paraformaldehyde at room temperature for 30 min. After removing fixing solution, the sections were washed 3 times with PBS and then incubated with blocking solution (goat sreum) for 1 h at room temperature. The sections were incubated with primary antibody and PIMO at 4 °C overnight. After wash, sections were incubated with corresponding secondary antibodies for 1 h at room temperature. Finally, sections were incubated with DAPI (Beyotime, China) for 5 min at room temperature, and then mounted with mounting medium containing anti-fluorescence quencher was added to mount the slides. Fluorescent images were captured using a confocal microscope (Zeiss, Germany).
Glycolysis metabolic measurements
PC cells were seeded on Seahorse XF24 culture cell plates at a concentration of 5.0 × 104 cells per well, and cells were incubated for 24 h under 20% or 1% O2. The probe plates were pretreated with 1 ml XF Calibrant in a non-CO2 incubator at 37 °C overnight. On the second day, the treatment media were removed and cultured cells were washed twice with assay medium Seahorse XF DMEM Medium (PH = 7.4) which is supplemented with 2 mM glutamine. Before detection, the culture cell plates were incubated for 1 h in a non-CO2 incubator at 37 °C. The final concentration of inhibitors and activators are 10 mM glucose, 1 μM Oligomycin, and 50 mM 2-DG. Cells were then assayed with an XFe24 extracellular flux analyzer (Seahorse XFe/XF). All the consumables were purchased from Agilent.
Statistical analysis
Data were analyzed with GraphPad Prism V9.0 and SPSS V19.0. The significance of differences between groups were assessed using Student’s t test or χ2 test as appropriate. The results are presented as mean ± standard error of mean (SEM) from three independent experiments. P value <0.05 was considered to be statistically significant. More information of the materials and methods is in the Supplementary Materials.
Data availability
All sequencing data generated in this study have been submitted to the NCBI Gene Expression Omnibus (GEO, https://www.ncbi.nlm.nih.gov/geo/) with accession number GSE218546.
References
**a T, Wu X, Cao M, Zhang P, Shi G, Zhang J, et al. The RNA m6A methyltransferase METTL3 promotes pancreatic cancer cell proliferation and invasion. Pathol Res Pract. 2019;215:152666.
Li BQ, Liang ZY, Seery S, Liu QF, You L, Zhang TP, et al. WT1 associated protein promotes metastasis and chemo-resistance to gemcitabine by stabilizing Fak mRNA in pancreatic cancer. Cancer Lett. 2019;451:48–57.
Gemenetzis G, Groot VP, Blair AB, Laheru DA, Zheng L, Narang AK, et al. Survival in Locally Advanced Pancreatic Cancer After Neoadjuvant Therapy and Surgical Resection. Ann Surg. 2019;270:340–7.
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71:209–49.
Melchionna R, Iapicca P, Di Modugno F, Trono P, Sperduti I, Fassan M, et al. The pattern of hMENA isoforms is regulated by TGF-beta1 in pancreatic cancer and may predict patient outcome. Oncoimmunology. 2016;5:e1221556.
Ashour AA, Abdel-Aziz AA, Mansour AM, Alpay SN, Huo L, Ozpolat B. Targeting elongation factor-2 kinase (eEF-2K) induces apoptosis in human pancreatic cancer cells. Apoptosis. 2014;19:241–58.
Tan Z, Xu J, Zhang B, Shi S, Yu X, Liang C. Hypoxia: a barricade to conquer the pancreatic cancer. Cell Mol Life Sci. 2020;77:3077–83.
**g X, Yang F, Shao C, Wei K, **e M, Shen H, et al. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol Cancer. 2019;18:157.
Zhang Y, Ren YJ, Guo LC, Ji C, Hu J, Zhang HH, et al. Nucleus accumbens-associated protein-1 promotes glycolysis and survival of hypoxic tumor cells via the HDAC4-HIF-1alpha axis. Oncogene. 2017;36:4171–81.
Roma-Rodrigues C, Mendes R, Baptista PV, Fernandes AR. Targeting Tumor Microenvironment for Cancer Therapy. Int J Mol Sci. 2019;20:840.
Kong F, Kong X, Du Y, Chen Y, Deng X, Zhu J, et al. STK33 Promotes Growth and Progression of Pancreatic Cancer as a Critical Downstream Mediator of HIF1alpha. Cancer Res. 2017;77:6851–62.
Lee HJ, Jung YH, Oh JY, Choi GE, Chae CW, Kim JS, et al. BICD1 mediates HIF1alpha nuclear translocation in mesenchymal stem cells during hypoxia adaptation. Cell Death Differ. 2019;26:1716–34.
Gu C, Wang Z, Zhou N, Li G, Kou Y, Luo Y, et al. Mettl14 inhibits bladder TIC self-renewal and bladder tumorigenesis through N(6)-methyladenosine of Notch1. Mol Cancer. 2019;18:168.
Li Q, Ni Y, Zhang L, Jiang R, Xu J, Yang H, et al. HIF-1alpha-induced expression of m6A reader YTHDF1 drives hypoxia-induced autophagy and malignancy of hepatocellular carcinoma by promoting ATG2A and ATG14 translation. Signal Transduct Target Ther. 2021;6:76.
Dong F, Qin X, Wang B, Li Q, Hu J, Cheng X, et al. ALKBH5 Facilitates Hypoxia-Induced Paraspeckle Assembly and IL8 Secretion to Generate an Immunosuppressive Tumor Microenvironment. Cancer Res. 2021;81:5876–88.
Chen Y, Peng C, Chen J, Chen D, Yang B, He B, et al. WTAP facilitates progression of hepatocellular carcinoma via m6A-HuR-dependent epigenetic silencing of ETS1. Mol Cancer. 2019;18:127.
He L, Li H, Wu A, Peng Y, Shu G, Yin G. Functions of N6-methyladenosine and its role in cancer. Mol Cancer. 2019;18:176.
Lobo J, Costa AL, Cantante M, Guimaraes R, Lopes P, Antunes L, et al. m(6)A RNA modification and its writer/reader VIRMA/YTHDF3 in testicular germ cell tumors: a role in seminoma phenotype maintenance. J Transl Med. 2019;17:79.
Fu Y, Zhuang X. m(6)A-binding YTHDF proteins promote stress granule formation. Nat Chem Biol. 2020;16:955–63.
Taketo K, Konno M, Asai A, Koseki J, Toratani M, Satoh T, et al. The epitranscriptome m6A writer METTL3 promotes chemo- and radioresistance in pancreatic cancer cells. Int J Oncol. 2018;52:621–9.
Oerum S, Meynier V, Catala M, Tisne C. A comprehensive review of m6A/m6Am RNA methyltransferase structures. Nucleic Acids Res. 2021;49:7239–55.
Geng H, Harvey CT, Pittsenbarger J, Liu Q, Beer TM, Xue C, et al. HDAC4 protein regulates HIF1alpha protein lysine acetylation and cancer cell response to hypoxia. J Biol Chem. 2011;286:38095–102.
Thalhammer A, Bencokova Z, Poole R, Loenarz C, Adam J, O’Flaherty L, et al. Human AlkB homologue 5 is a nuclear 2-oxoglutarate dependent oxygenase and a direct target of hypoxia-inducible factor 1alpha (HIF-1alpha). PLoS One. 2011;6:e16210.
Lan Q, Liu PY, Bell JL, Wang JY, Huttelmaier S, Zhang XD, et al. The Emerging Roles of RNA m(6)A Methylation and Demethylation as Critical Regulators of Tumorigenesis, Drug Sensitivity, and Resistance. Cancer Res. 2021;81:3431–40.
Mao Y, Dong L, Liu XM, Guo J, Ma H, Shen B, et al. m(6)A in mRNA coding regions promotes translation via the RNA helicase-containing YTHDC2. Nat Commun. 2019;10:5332.
Liu J, Dou X, Chen C, Chen C, Liu C, Xu MM, et al. N (6)-methyladenosine of chromosome-associated regulatory RNA regulates chromatin state and transcription. Science. 2020;367:580–6.
Yu J, She Y, Yang L, Zhuang M, Han P, Liu J, et al. The m(6) A Readers YTHDF1 and YTHDF2 Synergistically Control Cerebellar Parallel Fiber Growth by Regulating Local Translation of the Key Wnt5a Signaling Components in Axons. Adv Sci (Weinh). 2021;8:e2101329.
Wang X, Zhao BS, Roundtree IA, Lu Z, Han D, Ma H, et al. N(6)-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell. 2015;161:1388–99.
Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han D, et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature. 2014;505:117–20.
Huang H, Weng H, Sun W, Qin X, Shi H, Wu H, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20:285–95.
Wang YJ, Yang B, Lai Q, Shi JF, Peng JY, Zhang Y, et al. Reprogramming of m(6)A epitranscriptome is crucial for sha** of transcriptome and proteome in response to hypoxia. RNA Biol. 2021;18:131–43.
Aghdassi A, Sendler M, Guenther A, Mayerle J, Behn CO, Heidecke CD, et al. Recruitment of histone deacetylases HDAC1 and HDAC2 by the transcriptional repressor ZEB1 downregulates E-cadherin expression in pancreatic cancer. Gut. 2012;61:439–48.
Edderkaoui M, Chheda C, Soufi B, Zayou F, Hu RW, Ramanujan VK, et al. An Inhibitor of GSK3B and HDACs Kills Pancreatic Cancer Cells and Slows Pancreatic Tumor Growth and Metastasis in Mice. Gastroenterology. 2018;155:1985–98.e1985.
Yang J, Chheda C, Lim A, Hauptschein D, Zayou L, Tang J, et al. HDAC4 Mediates Smoking-Induced Pancreatic Cancer Metastasis. Pancreas. 2022;51:190–5.
Giaginis C, Damaskos C, Koutsounas I, Zizi-Serbetzoglou A, Tsoukalas N, Patsouris E, et al. Histone deacetylase (HDAC)-1, -2, -4 and -6 expression in human pancreatic adenocarcinoma: associations with clinicopathological parameters, tumor proliferative capacity and patients’ survival. BMC Gastroenterol. 2015;15:148.
Zhang X, Qi Z, Yin H, Yang G. Interaction between p53 and Ras signaling controls cisplatin resistance via HDAC4- and HIF-1alpha-mediated regulation of apoptosis and autophagy. Theranostics. 2019;9:1096–114.
**a X, Lemieux ME, Li W, Carroll JS, Brown M, Liu XS, et al. Integrative analysis of HIF binding and transactivation reveals its role in maintaining histone methylation homeostasis. Proc Natl Acad Sci USA 2009;106:4260–5.
Zhang C, Samanta D, Lu H, Bullen JW, Zhang H, Chen I, et al. Hypoxia induces the breast cancer stem cell phenotype by HIF-dependent and ALKBH5-mediated m(6)A-demethylation of NANOG mRNA. Proc Natl Acad Sci USA 2016;113:E2047–E2056.
Hou J, Zhang H, Liu J, Zhao Z, Wang J, Lu Z, et al. YTHDF2 reduction fuels inflammation and vascular abnormalization in hepatocellular carcinoma. Mol Cancer. 2019;18:163.
Wang Q, Chen C, Ding Q, Zhao Y, Wang Z, Chen J, et al. METTL3-mediated m(6)A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut. 2020;69:1193–205.
Pate KT, Stringari C, Sprowl-Tanio S, Wang K, TeSlaa T, Hoverter NP, et al. Wnt signaling directs a metabolic program of glycolysis and angiogenesis in colon cancer. EMBO J. 2014;33:1454–73.
Pavlova NN, Thompson CB. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016;23:27–47.
Ganapathy-Kanniappan S, Geschwind JF. Tumor glycolysis as a target for cancer therapy: progress and prospects. Mol Cancer. 2013;12:152.
Lin Z, Niu Y, Wan A, Chen D, Liang H, Chen X, et al. RNA m(6) A methylation regulates sorafenib resistance in liver cancer through FOXO3-mediated autophagy. EMBO J. 2020;39:e103181.
Jiang X, Liu B, Nie Z, Duan L, **ong Q, ** Z, et al. The role of m6A modification in the biological functions and diseases. Signal Transduct Target Ther. 2021;6:74.
Li Z, Peng Y, Li J, Chen Z, Chen F, Tu J, et al. N(6)-methyladenosine regulates glycolysis of cancer cells through PDK4. Nat Commun. 2020;11:2578.
Semenza GL. Hypoxia-inducible factors in physiology and medicine. Cell. 2012;148:399–408.
Keith B, Johnson RS, Simon MC. HIF1alpha and HIF2alpha: sibling rivalry in hypoxic tumour growth and progression. Nat Rev Cancer. 2011;12:9–22.
Tiwari A, Tashiro K, Dixit A, Soni A, Vogel K, Hall B, et al. Loss of HIF1A From Pancreatic Cancer Cells Increases Expression of PPP1R1B and Degradation of p53 to Promote Invasion and Metastasis. Gastroenterology. 2020;159:1882–97.e1885.
Ling S, Shan Q, Zhan Q, Ye Q, Liu P, Xu S, et al. USP22 promotes hypoxia-induced hepatocellular carcinoma stemness by a HIF1alpha/USP22 positive feedback loop upon TP53 inactivation. Gut. 2020;69:1322–34.
Lisy K, Peet DJ. Turn me on: regulating HIF transcriptional activity. Cell Death Differ. 2008;15:642–9.
Stegen S, Laperre K, Eelen G, Rinaldi G, Fraisl P, Torrekens S, et al. HIF-1alpha metabolically controls collagen synthesis and modification in chondrocytes. Nature. 2019;565:511–5.
Acknowledgements
This research was supported by grant from the Key Research and Development Program of Shandong Province (2019GHZ003, 2021ZLGX02 and 2020CXGC011304), Taishan Scholars Climbing Program of Shandong Province (NO.tspd20210323), Young Taishan Scholars Program of Shandong Province (NO.tsqn201909176), Outstanding Young and Middle-aged Scholar of Shandong University, Shandong Collaborative Innovation Center for R&D and Transformation of New Tumor Biomarkers Foundation (CXZX2019006), Tumor Biomarker Innovation Team Foundation of **an City (2019GXRC004 and 2021GXRC020), Shandong University Clinical Research Project (2020SDUCRCA002) and Natural Science Foundation of Shandong (ZR2020QH187).
Author information
Authors and Affiliations
Contributions
XL and MF completed the main experiments and data analysis. XH, ZG, ZW and YW provided some experimental materials and guidance. LD and CW wrote and modified the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics
The male C57 mice were raised in the SPF animal room of the Animal Center of the Second Hospital of Shandong University. All the animal experiments were performed following the ARRIVE reporting guidelines, and protocols were approved by the Institutional Animal Care and the Committee of the Second Hospital of Shandong University (approval ID: KYLL-2019(KJ)A-0264).
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, X., Feng, M., Hao, X. et al. m6A methylation regulates hypoxia-induced pancreatic cancer glycolytic metabolism through ALKBH5-HDAC4-HIF1α positive feedback loop. Oncogene 42, 2047–2060 (2023). https://doi.org/10.1038/s41388-023-02704-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41388-023-02704-8
- Springer Nature Limited
This article is cited by
-
N6-methyladenosine demethyltransferase FTO mediated m6A modification of estrogen receptor alpha in non-small cell lung cancer tumorigenesis
Oncogene (2024)
-
RNA modifications in the progression of liver diseases: from fatty liver to cancer
Science China Life Sciences (2024)
-
Mechanisms and clinical landscape of N6-methyladenosine (m6A) RNA modification in gastrointestinal tract cancers
Molecular and Cellular Biochemistry (2024)