Abstract
Pax6 is a highly conserved transcription factor among vertebrates and is important in various aspects of the central nervous system development. However, the gene regulatory circuitry of Pax6 underlying these functions remains elusive. We find that Pax6 targets a large number of promoters in neural progenitors cells. Intriguingly, many of these sites are also bound by another progenitor factor, Sox2, which cooperates with Pax6 in gene regulation. A combinatorial analysis of Pax6-binding data set with transcriptome changes in Pax6-deficient neural progenitors reveals a dual role for Pax6, in which it activates the neuronal (ectodermal) genes while concurrently represses the mesodermal and endodermal genes, thereby ensuring the unidirectionality of lineage commitment towards neuronal differentiation. Furthermore, Pax6 is critical for inducing activity of transcription factors that elicit neurogenesis and repress others that promote non-neuronal lineages. In addition to many established downstream effectors, Pax6 directly binds and activates a number of genes that are specifically expressed in neural progenitors but have not been previously implicated in neurogenesis. The in utero knockdown of one such gene, Ift74, during brain development impairs polarity and migration of newborn neurons. These findings demonstrate new aspects of the gene regulatory circuitry of Pax6, revealing how it functions to control neuronal development at multiple levels to ensure unidirectionality and proper execution of the neurogenic program.
Similar content being viewed by others

Introduction
The paired box protein, Pax6, is a highly conserved transcription factor of 422 amino acids comprising two DNA-binding domains, an amino-terminal paired domain and a homeodomain along with a carboxy-terminal proline/serine/threonine-rich transactivation domain [1, 2]. Pax6 was first discovered to be required for proper segmentation in Drosophila [3, 4] and later shown to be essential for eye development in Drosophila [5], a role that was further found to be conserved in human and mouse eye development [6, 7]. During mammalian brain development, Pax6 is expressed in a specific spatiotemporal manner and is restricted to mainly neuronal tissues [2, 8]. Pax6 is now established to be essential for maintaining the pool of neural stem cells (NSCs) and thereby regulating embryonic as well as adult neurogenesis, as shown by its expression in neuroepithelial and radial glial cells, which can divide symmetrically to produce NSCs or asymmetrically to become a NSC and a neuron [9, 10].
The discovery of a plethora of known Pax6 functions has been facilitated by various Pax6 mutants. One such very useful mutant, the small eye (Sey) mouse mutant, contains a single-base substitution [11], resulting in the production of a functionally inactive and truncated Pax6 lacking the DNA-binding homeodomain and the C-terminal activation domain. Importantly, the Sey mutant mouse phenotype is similar to that of an artificially targeted Pax6-deficient mouse (Pax6−ax), showing small eyes and numerous neural defects, including reduced neurons in the cerebral cortex [11–13]. These phenotypic similarities in the Sey mutant and Pax6−ax substantiate the use of Sey homozygous mutant mice as Pax6-null mutants. It was further shown that Sey mutant embryonic stem (ES) cells generate misspecified neurons that undergo death because of high expression of the neurotrophin receptor p75NTR [14].
It is well established that Pax6 is crucial for the development of the central nervous system, eyes, nose, pancreas and pituitary gland [13, 15, 16]. Recent studies have shown that Pax6 functions upstream of gene networks involved in brain patterning, neuronal migration and neural circuit formation [17]. Despite the established role of Pax6 in neurogenesis, its genomic targets, their chromatin status and its cooperativity with other transcription factors during neurogenesis remain unclear. Furthermore, while a number of players functioning downstream of Pax6 have been identified, these are not enough to explain the plethora of functions Pax6 is known for. Here we reveal that in neural progenitors, Pax6 binds a large number of gene promoters that exhibit epigenetic state that is hallmark of open chromatin. Many Pax6-bound promoters are also targeted by Sox2 and functionally cooperate in gene regulation underlying neuronal specification. Pax6 directly binds and silences genes important for mesoderm and endoderm development as they get de-repressed in progenitors lacking Pax6. In addition, Pax6 targets that are downregulated in mutant progenitors are known to be critically involved in neuronal development. Pax6-driven gene-expression program further induces activity of neurogenic transcription factors and repress others that promote non-neuronal lineages. Importantly, our analysis also revealed a number of Pax6-induced genes that are highly expressed during brain development but their function has not yet been explored during neurogenesis. Here we show that one such gene, Ift74, which is directly bound and activated by Pax6 in NP cells, is required for the proper migration of newborn neurons. Furthermore, our analysis revealed that Pax6 directly targets the promoter of Notch signaling components and induces their expression, which then further contribute to Ift74 expression. These observations reveal the manner by which Pax6 controls multiple components of the network underlying neuronal development and uncovers Ift74 as a novel regulator of neurogenesis.
Results
Pax6 binds to a number of gene promoters in neural progenitors cells
We first determined the expression patterns of Pax6 in various embryonic tissues and cortical layers. As shown previously, Pax6 is specifically highly expressed in ventricular zone (VZ) and is gradually lost as cells progress through the subventricular zone (SVZ) to the cortical plate (CP; Supplementary Figure S1A). An analysis of other ectoderm (epidermis), mesoderm (heart and mouse embryonic fibroblasts) and endoderm tissues (lung and pancreas) showed relative absence of Pax6 expression, with the exception of the pancreas that exhibited low levels of Pax6, confirming previous reports [18] (Supplementary Figure S1A). We next use a highly refined and established differentiation model of neurogenesis, in which mouse ES cells first differentiate into Pax6-positive NP (radial glial-like) cells (also referred as celllular aggregates, in short CA_D8) and subsequently into terminally differentiated glutamatergic pyramidal neurons (TN) with high purity (>95%) and synchrony and is known to closely recapitulate the stages of embryonic neurogenesis [19–21]. The expression analysis of Pax6 in this system revealed its highest expression in cellular aggregate cells, thereby presenting a system for investigating Pax6 function in vitro (Supplementary Figure S1B).
To shed light on Pax6 function, we performed Pax6 chromatin immunoprecipitation (ChIP) in NPs and investigated its genome-wide binding pattern using a previously described ChIP-chip platform in biological replicates [22]. These arrays cover 10% of the mouse genome, including all well-annotated promoters, several large multigene loci and the complete chromosome 19 [23]. The visual inspection of the genomic regions suggested that Pax6 is targeted to distinct genomic sites and also occupies a number of promoters (n=5086, promoter enrichment >0.25; Figure 1a and b, Supplementary Figure S1C and D, Supplementary Table S1). A comprehensive and unbiased analysis of Pax6 binding along the fully tiled chromosome 19 revealed its relatively high enrichment at promoters (Figure 1c). These observations were validated at selected gene promoters in independent ChIP assays (Figure 1d). Such targeting of Pax6 to gene promoters prompted us to investigate its relationship with the chromatin state of target sites and the transcriptional states of associated genes at the progenitor stage. We analyzed the ChIP-seq data sets for RNA Pol II, H3K4me2, which is an established active histone modification, and the Polycomb group repressive mark, H3K27me3, at the NP stage in the same differentiation system, and correlated these data with Pax6 occupancy at the target gene promoters (Figure 1e and f). Further analysis showed that Pax6 occupancy was most strongly correlated with the active mark H3K4me2 (R2=0.64; Figure 1e). In addition, a large number of Pax6-bound promoters were RNA Pol II bound (R2=0.44) and actively transcribed (R2=0.57; Figure 1e). Furthermore, Pax6 target promoters were mostly devoid of the repressive mark H3K27me3 (R2=0.13; Figure 1e). Heat map visualization at promoters supported these observations, revealing that the majority of Pax6-bound genes displayed the H3K4me2 mark, a significant fraction of which were Pol II bound and actively transcribed (Figure 1f). Furthermore, comparison of Pax6 occupancy at promoters with enrichment of RNA Pol II, H3K4me2 and H3K27me3 in the same differentiation system showed similar patterns (Figure 1g). A comparison with promoter targets of Pax6 recently identified by ChIP-seq assay in E12.5 forebrain tissue [34–36] (Supplementary Figure S3C and D). Although previously Pax6 has been indirectly implicated in the control of Notch pathway [10], our analysis revealed that Pax6 directly binds at the promoters of a large number of genes associated with Notch signaling. Furthermore, these genes were downregulated in mutant NPs, indicating that Pax6 has a direct role in the activation of Notch signaling in NP cells (Figure 3j). This targeting by Pax6 at Notch signaling components provides potential mechanism regarding how this master transcription factor acts at multiple levels to define progenitor identity and differentiation towards neurons. Overall, these analyses identify a dual role for Pax6, in which it mediates the activation of neuronal (ectodermal) genes while concurrently represses the mesodermal and endodermal genes.
Pax6 influences transcription factor network to confer unidirectionality towards neuronal differentiation
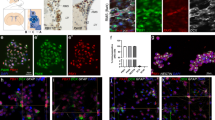
On the basis of our observations that Pax6 mutant cells showed upregulation of non-neuronal and downregulation of neuronal genes, we next probed whether activities of any particular transcriptional factors are altered in the absence of Pax6 that in turns could explain part of gene-expression program alterations. Towards this we applied integrated system for motif activity response analysis (ISMARA), which predicts the transcription factors that can potentially regulate the differentially expressed genes on the basis of binding motifs at the promoters of these genes [37,38]. ISMARA analysis predicted a number of transcription factors whose activity significantly changed in Pax6 mutant NPs. This included Pax6 and Sox2 that showed downregulation in their activity (Figure 4a–f). In line with these findings, ISMARA predicted targets of Pax6 and Sox2 were also found to be significantly downregulated in Sey cells (Figure 4b and e). Furthermore, ISMARA also predicted interaction networks of Pax6 and Sox2 with other transcription factors many of which are known to be important for neurogenesis (Figure 4c and f). Two transcription factors, TFAP2B and TCF4, were commonly identified in both Pax6 and Sox2 interaction networks (Figure 4c, f and g). Surprisingly, predicted targets of TFAP2B or TCF4 were highly enriched for genes related to neurogenesis (Supplementary Table S3). The role of TFAP2B in neuronal development as well as its interaction with Pax6 and Sox2 is unknown, however, our prediction provides potential insights of how cooperativity between different transcription factors contributes to neurogenesis. Targets of TFAP2B were also very significantly downregulated in Sey cells (Figure 4h) and network analysis further predicted its interaction with Pax6 as well as Sox2 in addition to many other interesting factors known to be required for neurogenesis (for example, Zeb1; Figure 4i). Overall, these observations suggest a cooperative function of transcription factors Pax6, Sox2 and TFAP2B in WT progenitors in gene activation as their activity and consequently their targets are downregulated in mutant cells.

Pax6 is critical for inducing activity of transcription factors that elicit neurogenesis and repress others that promote non-neuronal lineages. (a) Pax6 motif identified by ISMARA. (b) Box plot showing expression in WT and mutant of Pax6 targets predicted by ISMARA. (c) First-level interaction network of Pax6 and its potential targets as predicted by ISMARA. (d) Sox2 motif identified by ISMARA. (e) Box plot showing expression in WT and mutant of Sox2 targets predicted by ISMARA. (f) First-level interaction network of Sox2 and its potential targets as predicted by ISMARA. (g) Tfap2b motif identified by ISMARA. (h) Box plot showing expression in WT and mutant of Tfap2b targets predicted by ISMARA. (i) First-level interaction network of Tfap2B and its potential targets as predicted by ISMARA. (j) Brachyury (T) motif identified by ISMARA. (k) Box plot showing expression in WT and mutant of Brachyury (T) targets predicted by ISMARA. (l) First-level interaction network of Brachyury (T) and its potential targets as predicted by ISMARA. (m) Myf family motif identified by ISMARA. (n) Box plot showing expression in WT and mutant of Myf family targets predicted by ISMARA. (o) First-level interaction network of Myf family and its potential targets as predicted by ISMARA. (p) Hnf1a motif identified by ISMARA. (q) Box plot showing expression in WT and mutant of Hnf1a targets predicted by ISMARA. (r) First-level interaction network of Hnf1a and its potential targets as predicted by ISMARA. All P-values are calculated using Wilcoxon test.
Furthermore, ISMARA analysis also revealed upregulation in the activity of a number of transcription factors that are known to be important for non-neuronal lineages such as T (brachyury), Hnf1a and members of the Myf family (Figure 4j–r). Brachyury is an established mesoderm transcription factor [37,38], while Hnf1a is critical for liver differentiation [39] and Myf family of transcription factors are known to be crucial for heart development [37]. Target genes of these three transcription factors were significantly upregulated in mutant cells (Figure 4k, n and q). GO enrichment analysis showed that these target genes are involved in the development and function of non-neuronal tissues (Supplementary Table S3). Furthermore, the network for each of these factors mostly consisted of a non-overlap** set of transcription factors (Figure 4l, o and r). Overall, these findings show that Pax6-dependent gene regulatory circuitry induces activity of transcription factors that induce neurogenesis and repress others that promote non-neuronal lineage.
Sox2 targets a large number of Pax6-bound gene promoters
We were intrigued by our observations that Sox2 activity is significantly reduced in Pax6 mutant NP cells. Both Sox2 and Pax6 are known to be important for the maintenance of the proliferative and developmental potential of NSCs [40]. Although it is known that Pax6 and Sox2 form a complex [41, 42], it remains to be investigated whether they function together in gene regulation at the same targets sites in the genome. We, therefore, compared our list of Pax6 target promoters with that of Sox2-bound promoters in NP cells derived from mouse ES cells in a previous study [43]. This analysis revealed that both Pax6 and Sox2 co-occupy a noticeable set of gene promoters, suggesting a potential cooperativity between these two transcription factors in gene regulation (Figure 5a and Supplementary Figure S4A; hypergeometric P-value: 1.28e-65). We next classified the genes encoding transcription factors, which were either expressed or repressed in the NPs in vivo (based on the transcriptome analysis of the E14.5 VZ cells) [Full size image
To further delineate and substantiate the expression dynamics of Pax6 and Sox2 targets, we explored recently published transcriptome data sets for distinct progenitor subpopulations (aRG, apical radial glial; bRG, basal radial glial; IPC, intermediate progenitors) as well as neurons from develo** mouse neocortex [44]. Comparison of NP markers in our in vitro neuronal differentiation system and the above data sets showed that our ES-derived progenitors are apical in nature (Supplementary Figure S4C and D). Further comparison revealed an interesting pattern of expression for Pax6 only and Sox2 only bound genes compared with Pax6 and Sox2 co-occupied genes during neurogenesis (Figure 5f and h). The set of genes bound by either Pax6 or Sox2 and expressed in aRG were repressed in the immediate next stage (bRG) and remained repressed throughout neurogenesis (cluster A in Figure 5f and g). However, the genes bound by either Pax6 or Sox2 and repressed in aRG showed transcriptional activation in a stage-specific manner during neurogenesis (cluster B, C and D Figure 5f and g). In contrast, genes co-occupied by both transcription factors and expressed (cluster A) or repressed (cluster B) in aRG were immediately repressed or activated in bRG, respectively, and maintained this state throughout neurogenesis (Figure 5h). Overall, these observations suggest that the gene regulatory function of Pax6 at its target sites may be influenced by co-factors such as Sox2.
We next wondered whether other transcription factors expressed later during neurogenesis could function at Pax6 and Sox2 target sites when Pax6 and Sox2 are no longer available. To test this hypothesis we chose Ascl1, which is shown to be essential for the transition from neuronal progenitors to a neuronal state [45, 46] and neurogenesis is severely impaired in the absence of Ascl1 [47–49]. During neuronal differentiation from ES cells, Pax6 and Sox2 are simultaneously highly expressed in NP cells and following onset of neurogenesis, their levels decrease while Ascl1 levels are further increased (Supplementary Figures S4E). Using a recently published genome-wide binding data set for Ascl1 during neurogenesis [50], we found that Ascl1 shared 44% of Pax6 (hypergeometric P-value, 1.55e-22) and 25% (hypergeometric P-value, 3.24e-13) of Sox2 targets (Figure 5i and j). Interestingly, the targets common between Pax6, Sox2 and Ascl1 (n=75) included classical Notch pathway (Id1, Id2, Hey1, Hes6 and Dll1) and neuronal (Tubb2b, Robo1, Mapt and Pcdh10) genes. We next explored how Pax6 and Sox2 targets that are also bound by Ascl1 are expressed during in vitro neurogenesis. Heat map visualization of these sets showed that such genes that are bound by Pax6 and/or Sox2 and also by Ascl1 mostly maintain their transcription state as cells exit NP state (higher Pax6/Sox2 and lower Ascl1 levels) towards initiating neurogenesis (lower Pax6/Sox2 and higher Ascl1 levels; Figure 5k and m). Furthermore, most of these genes acquire an opposite expression state in terminally differentiated neurons (no Pax6, Sox2 or Ascl1 expression). This suggests that a distinct sets of Pax6/Sox2 target genes might be targeted by other transcription factors in subsequent stages of neurogenesis to facilitate maintenance of their transcription state despite the later absence of Pax6/Sox2 itself.
To further explore the functional differences between the genes occupied by Ascl1 uniquely or Ascl1 along with Pax6 and/or Sox2, we performed a comparative GO term analysis to reveal their possible involvement in any specific biological processes (Figure 5n). The set of genes that were targeted by Pax6 and Sox2 only (blue squares) were enriched for a broad range of functions related to neural precursor or neural tube formation, cell cycle, transcription regulation, protein localization, metabolic processes and chromatin organization (Figure 5n). These genes were also enriched for functions related to neuronal differentiation and maturation (yellow squares). Interestingly, Ascl1 unique target genes were also enriched for these functions (yellow squares) indicating towards a functional takeover of neuronal development by Ascl1 (Figure 5n). The set of genes that were bound by Pax6/Sox2 complex and also by Ascl1 were exclusively enriched for Notch signaling and neuronal differentiation (Figure 5n, red squares). We also observed that the functional class ‘neuronal projection’ was uniquely attributed to targets that were also targeted by Ascl1 only (green squares; Figure 5n), supporting its known role in early neuronal development. Overall these data indicate that a subset of neurogenesis-related genes that are acted upon by Pax6 and/or Sox2 in NPs may also be targeted by other transcription factors such as Ascl1 for gene regulation during neuronal development.
Pax6 directly induces expression of many known and novel NP-specific transcription factors
We next attempted to further investigate the role of Pax6 in regulating the expression of NP-specific genes by performing a series of stepwise analyses. First, we selected the Pax6-bound mutant-downregulated genes that were significantly higher expressed in E14.5 cortical layers compared with other tissues (heart, embryonic fibroblasts, lung and pancreas). Then, we selected those factors that were at least two-fold upregulated in the VZ compared with the CP (Figure 6a). Interestingly, this final list of 46 genes primarily consisted of transcription factors, including established Pax6 targets and known regulators of NP identity (for example, Nestin, Neurog1/2, Neurod1/4 and Notch pathway components, such as Dll1 and Hes6; Figure 6b and Supplementary Figure S5A). Pax6 was also bound to its own locus likely for autoregulation as shown previously [12]. Of these 46 Pax6-target gene promoters, 17 were also co-occupied by Sox2 (data not shown). This analysis also identified many novel factors that have not been previously shown to function in regulating progenitor identity (Supplementary Figure S5B). The expression pattern of many of these genes was further validated by their in situ hybridization analysis in the embryonic cortex (Figure 6c) [51]. This analysis revealed how Pax6 functions as an upstream regulator of many known critical neurogenesis-related transcription factors, at the same time identified many previously unknown Pax6 targets that are specifically expressed in the cortex and warrant further investigation.

Pax6 directly induces expression of a large number of known and novel neural progenitor-specific transcription factors. (a) Flow chart showing the identification of genes that are specifically upregulated in neural progenitors in vivo and are regulated by Pax6. (b) Heat map showing the expression patterns of 46 genes identified in a. Red means high expression while blue means lower expression. (MEF, mouse embryonic fibroblast). (c) In situ hybridization images for known and novel Pax6 targets as derived from the Allen Brain Atlas (http://develo**mouse.brain-map.org/).
Ift74 is a novel Pax6 target that contributes to neuronal migration
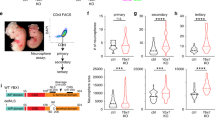
We were next interested to deeply explore the function of novel genes that were directly bound and activated by Pax6 and whose expression was restricted to NPs. We focused on Ift74 (intraflagellar transport (IFT) 74 homolog), which is a component of the IFT complex but remains a rather uncharacterised protein in the context of mammalian biology. Ift74 forms a tubulin-binding module together with IFT81 that specifically mediates transport of tubulin within the cilium required for ciliogenesis [52]. To precisely map the kinetics of its expression with respect to Pax6, we analyzed their expression at various time points during the differentiation of ES cells into neurons via a NP state. As expected, this fine time course analysis during neuronal differentiation revealed that Pax6 was most highly induced upon commitment to NPs and downregulated as soon as neurogenesis progressed (Figure 7a). Interestingly, analysis of Ift74 at same time points showed that it reached its maximum expression levels few hours after highest Pax6 expression, a stage that marks the transition of NP cells to neurons, and subsequently its expression was reduced upon neuronal maturation (Figure 7a).

Ift74 contributes to neuronal migration. (a) Expression of Pax6 and Ift74 during neuronal differentiation of ES cells derived by real time quantitative PCR (RT-qPCR). Expression is shown for various stages of neuronal differentiation (ES cells, CA at day 4 before adding RA (CA d4), CA at day 8 (CA_D8)) and various time points during neurogenesis (TN at 12 h and day 1, 2, 3, 5, 7 and 10). mRNA expression is normalized to the housekee** gene Rpl19 (n=3, error bars show S.E.M). (b) Expression of Ift74 (in RPKM) in VZ, SVZ, and CP dissected from mouse embryos at E14.5 derived by RNA-sequencing (GSE30765). (c) Pax6 ChIP-qPCRs to validate Pax6 binding at the promoter (−200±0 bp region) of Ift74 at CA day 8 (CA_d8) (n=3, error bars shown as s.e.m.). Average enrichments are plotted normalized to input and further to an intergenic control region (control). (d) Fold change in mRNA levels in CA_d8 cells treated with γ-secretase inhibitor as compared with non-treated cells. ES cells were induced to undergo neuronal differentiation and treated every other day with 5 μM N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) (n=5), 3 nM LY-411575 (n=3) or dimethylsulfoxide as control from CA_d4 stage onwards. Expression of the shown genes were normalized to Rpl19 levels (ΔCT) and fold change with respect to the control is plotted (error bars show s.e.m.). Statistical significance were calculated with an unpaired t-test (*P<0.05; **P<0.01; ***P<0.001; ****P<0.0001). (e) Left panel: representative immunofluorescence images from coronal brain sections at E16.5 stained for DNA using Hoechst (blue). Green fluorescent protein (GFP; green) marks shIft74 or non-targeting control electroporated cells; brains were electroporated at E12.5 and analyzed after 4 days. Scale bar: 100 μm. Right panel: quantifications of fluorescence signal in the GFP channel using ImageJ in the lower Hoechst dense region (VZ/SVZ), the intermediate less DNA dense region (IZ) and the upper Hoechst dense region (CP). Error bars reflect s.e.m. of three representative regions from two independently electroporated brains. (f) Representative immunofluorescence images of cortical brain slices electroporated at E12.5 with non-targeting control or shIft74 and analyzed at E16.5. GFP (green) and PAX6 (red) stain is represented and the scale bar: 100 μm. All GFP-positive cells have been counted and the percentage of cells that also displayed PAX6 signal is plotted on the y axis n=2, error bars show s.e.m.). (g) Same as in f but co-stained with TBR2 (red) and accordingly quantified as above for TBR2-positive electroporated cells. (h) Representative immunofluorescence images of cortical brain slices electroporated at E12.5 with shIft74 and analyzed at E16.5. GFP (green), TUJ1 (red) and DNA (blue) stain is represented and the scale bar: 100 μm. (i–k) GFP images to analyze cell shape and polarity for non-targeting control and shIft74 electroporated brains in VZ/SVZ (i), IZ (j) and CP (k) region. White arrowheads point to multipolar roundish cells in the IZ and bipolar cells in VZ, SVZ and CP of non-targeting control and shIft74 electroporated brains. Scale bar: 50 μm. On the right side of each image representative thresholded cells of the particular layer are represented.
To further substantiate our observations of Ift74 induction in the context of Pax6 expression in vivo, we next analyzed transcriptome data derived from the three layers of the E14.5 cortex (VZ, SVZ and CP) that showed the prominent expression of Ift74 in the VZ of the develo** mouse brain [63]. In this study, we attempted to uncover genes under the transcriptional control of Pax6 in NPs and identified downstream transcription factors that contribute to neurogenesis. We found that in NPs, Pax6 is targeted to many promoters that showed a distinct epigenetic state of open chromatin. Interestingly, many Pax6 sites are also occupied by Sox2, suggesting that they function together in gene regulation. Pax6 deficiency causes defects in the expression of its target genes, linking its binding to a function in transcriptional regulation. Strikingly, our analysis also revealed a dual role for Pax6, in which it activates the neuronal (ectodermal) genes while concurrently represses the mesodermal and endodermal genes. Importantly, Pax6 also directly induces the expression of a number of known as well as novel genes including transcription factors that are specifically expressed in NPs. We further show that one of the novel Pax6 target gene, Ift74, may have an important role during neurogenesis, likely via regulating migration of newborn neurons. Furthermore, our results also provide indication that Notch signaling contributes to the transcriptional induction of Ift74 in NPs. Interestingly, Pax6 also directly binds at the promoters of many Notch signaling components and functions in their activation, suggesting the functional cooperativity between Pax6 and Notch signaling in regulating a downstream gene-expression program. Overall, our findings reveal how Pax6 regulates the gene-expression program at multiple levels to ensure proper execution of the neurogenic program, and at the same time ensures the unidirectionality of neuronal differentiation (Figure 8).

Pax6 regulates the gene-expression program at multiple levels to promote neuronal differentiation. Pax6 mediates the activation of neuronal (ectodermal) genes while concurrently represses the mesodermal and endodermal genes, thereby ensuring the unidirectionality of lineage commitment towards neuronal differentiation. Pax6 directly binds and activates expression of critical transcription factors and components of signaling pathways, all of which then function in concert to orchestrate downstream gene-expression program that drives neurogenesis.
The complexity of Pax6 function has been suggested to arise from its interaction with various transcription factors to synergistically regulate target gene expression. In lens development, the transcriptional regulation of several crystallin genes by Pax6 is achieved in coordination with other transcription factors, such as Sox2 and Maf [64, 65]. Pax6 has been shown to form a complex with Sox2 to transcriptionally activate the δ- c rystallin gene [41]. Sox2 is of further relevance because it is expressed in the develo** mouse central nervous system from an early stage [66] and regulates the expression of fibroblast growth factor 4 (Fgf4) and Nestin, which are important in maintaining NSCs [67]. Our ISMARA analysis showed that the activity of Sox2 was significantly reduced in Pax6 mutant NP cells. Moreover, Sox2 co-occurred with Pax6 at many promoters in NP cells. Furthermore, these data also reveal that critical NP genes, such as Nestin, are co-regulated by Pax6 and Sox2. We also observed that the target genes co-occupied by Pax6 and Sox2 are expressed at higher levels, including those encoding important transcription factors, as compared with Pax6 only targets. These findings highlight the importance of the interplay between Pax6 and Sox2 in cooperative transcriptional regulation during neurogenesis, and at the same time also imply that critical neurogenesis genes may require co-activation by more than one stage-specific transcription factor. Furthermore, these observations also indicate that the gene regulatory potential of Pax6 may be determined by its partners and in this specific case, Sox2 occupancy drives it more towards a transcription activating role.
We also observed distinct expression dynamics of Pax6 and Sox2 unique target genes compared with those co-occupied by both factors during subsequent stages of neurogenesis. The genes bound by either Pax6 or Sox2 and expressed in aRG showed transcriptional activation in a stage-specific manner during neurogenesis where different gene-sets were found to be expressed in bRG, IPC and neurons. It may reflect availability of distinct factors or signaling pathways that become active at each of these stages to induce a set of genes critical for that particular stage of neuronal differentiation. On the other hand, all genes co-occupied by both Pax6 and Sox2 acquired changes in their expression state immediately after transition from aRG to bRG and this transcription state was maintained during later stages. This may also imply that genes that are required to be immediately switched on or off during differentiation of NPs may in some way benefit from being targeted by both Pax6 and Sox2.
Pax6 is believed to exert its effects by regulating critical downstream effectors during neurogenesis. A number of such examples have already been described, such as Fabp7, Neurog2, p27kip1, cell adhesion molecules (for example, L1 optimedin A, R-cadherin, δ-catenin and tenascin C), patterning molecules (for example, secreted frizzled-related protein 2 (sFRP2) and T-cell factor 4 (Tcf4)), Nkx2.2, Hoxd4, as well as other transcription factors, including Nfia, AP-2γ, NeuroD6, Neurog2, Tbr2, and Bhlhb5 [9, 15, 16, 57–63, 68, 69]. Our study is in line with the previous reports of a direct regulation of neurogenic transcription factors by Pax6 [17]. Importantly, our data also identified several additional genes including transcription factors that are directly induced by Pax6 in NP cells but have not been studied in the context of neuronal development (for example, Bazb2, Hmgn3, Peli2, Vit). Furthermore, although the role of Pax6 and Notch signaling in neuronal development is known for long, our data provide the first evidence that Pax6 also promotes Notch signaling by directly inducing the expression of critical components of this pathway (Figure 8).
Our data suggest that although Pax6 activates genes related to neuronal development, it represses the transcription of mesodermal and endodermal genes. Furthermore, ISMARA analysis also showed that Pax6 is required for the induction of transcription factors and their targets that elicit neurogenesis (for example, Sox2 and Tfap2b) and repress others that promote non-neuronal lineages (for example, Brachyury, Hnf1a and Myf family of transcription factors). These observations strongly imply that Pax6-driven gene regulatory program functions to ensure the unidirectionality towards neuronal differentiation.
During neurogenesis, NP cells undergo massive morphological and spatial changes that are tightly linked to cytoskeleton changes. For example, neocortical neurons arise by asymmetric division of radial glia progenitors (RG) in the VZ with a bipolar morphology and gradually become multipolar as they reach SVZ/IZ zone and move erratically. Subsequently, these cells undergo a multipolar to bipolar transition and move rapidly along RGs to the top of CP [70]. Defective ciliogenesis has been shown to accompany defects in neuronal migration in human ciliopathy phenotypes such as Meckel–Gruber syndrome [71]. We find that one of the Pax6-induced genes, Ift74, is highly expressed in NP cells as compared with other cell types. It has been shown in human cells that Ift74 and Ift81 build a tubulin-binding module whose binding to tubulin is important for ciliogenesis [52]. Depletion of Ift74 by in utero electroporation during cortical development led to a retention of migrating cells in the lower layer of the cortex. Furthermore, some cells showed multiple small processes, indicating that the knockdown cells are in the multipolar phase in SVZ/IZ region and fail to migrate to the CP. These observations collectively suggest that the regulation of ciliogenesis and/or axonogenesis via Ift74 might be essential for cortical development. Moreover, Ift74-depleted cells showed significantly reduced migration capacity during EMT in vitro. Importantly, as Notch signaling is known to be essential for proper radial migration of cortical neurons [72] and since we also found that Notch signaling is required for proper transcriptional induction of Ift74, it is likely that the previously observed defects in neuronal migration in the absence of Notch signaling are, at least in part, contributed by a loss of Ift74 expression.
Taken together our findings provide novel insights into genomic localization and gene regulatory function of Pax6 during cortical development. Here we show that Pax6 targets a distinct class of epigenetically marked gene promoters, a number of which are co-occupied by other critical transcription factors such as Sox2. Our results suggest a model for a dual function of Pax6 upon neuronal commitment where it mediates the activation of neuronal (ectodermal) genes while concurrently represses the mesodermal and endodermal genes to ensure the unidirectionality towards neuronal differentiation. In addition, Pax6 also induces critical signaling pathways that further work together with Pax6 in guiding critical neurogenic events. Our findings highlight how the gene regulatory circuitry organized by a single factor is able to contribute to neuronal development at multiple levels. In addition to many established downstream effectors, this study has identified many novel targets that are bound and activated by Pax6 and warrant further investigation in cortical development. The in utero knockdown of one such gene, Ift74, during brain development resulted in impaired neuronal polarity and migration of newborn neurons. Overall, these findings reveal how Pax6 functions in the control of neuronal development at multiple levels to ensure unidirectionality and proper execution of the neurogenic program.
