
Abstract
One of the most remarkable aspects of Devil Facial Tumour Disease (DFTD) is its infectious nature, and for successful transmission it must avoid detection by the devil’s immune system. For this to occur, the devil either is severely immunosuppressed or factors produced by the tumor contribute to its avoidance of immune detection. An analysis of the devil’s immune system revealed the presence of normal-looking lymphoid organs and lymphoid cells. At a functional level the lymphocytes proliferated in response to mitogen stimulation. Subcutaneous injection of a cellular antigen produced a strong antibody response, providing compelling evidence that the devil has a competent immune system. Tumor cell analysis demonstrated that the tumor expresses the genes of the major histocompatibility complex; however, there was a limited diversity. Therefore, the most likely explanation for devil-to-devil transmission of DFTD is that the tumor is not recognized by the devil as “non-self” because of the limited genetic diversity. With its consistent morphology and relatively stable genome, this tumor would provide a reasonable target for a vaccine approach, provided the immune system can be coaxed into recognizing the tumor as “non-self.”
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Devil Facial Tumour Disease (DFTD) and canine transmissible venereal tumor are both unique wildlife diseases because they are tumors that are directly transferred between unrelated animals (Das and Das, 2000; Murgia et al., 2006; Pearse and Swift, 2006). One particularly unique aspect of DFTD is that there is no evidence of host resistance. The tumor is malignant (i.e., the primary tumor grows rapidly with abundant mitotic indexes and it frequently metastasizes) and, as understood to date, “infection” by this tumor results in the death of the animal (Loh et al., 2006a).
That DFTD is transferable between unrelated devils was confirmed by a karyotypic analysis of tumor versus host cells (Pearse and Swift, 2006). Because the tumor displays a different chromosomal arrangement than that of the host and because all tumors are highly likely to have been derived from the same rogue cell line, the tumor appears to be transferred in a manner analogous to an organ transplant between unrelated animals. Under normal circumstances, when any organ or tissue, including cancer, is transplanted from one host to another, it is rejected by the immune system of the recipient. Organ transplants in humans are successful only by carefully matching immune response genes, known as the MHC (major histocompatibility complex), between the transplanted organ and the recipient. Even then, closely matched transplants are rejected unless the recipient receives therapy to suppress their immune system. Identical MHC matches (e.g., between identical twins) are the exception that do not require immunosuppression. With the ease of transfer of DFTD between unrelated devils, the status of their immune system must be questioned and consequently immune investigations are clearly warranted. In this article we present and review our data on in vitro and in vivo assessments of the immune response of healthy disease-free Tasmanian devils to provide insight into why otherwise healthy normal animals should succumb to an “infectious cancer.”
Methods
Tasmanian Devils
Tasmanian devils kept in captivity by the Department of Primary Industries and Water (DPIW) were used as the source of normal blood samples. All devils were anesthetized in an open system with isofluorane delivered via mask and 10-20 ml of blood was collected from the jugular vein. Blood was immediately transferred to tubes containing lithium heparin and kept at room temperature until arrival in the laboratory.
Animals for autopsies and for normal lymphoid tissues were healthy devils that were obtained when recent road kills became available or when severely injured devils were euthanized by qualified veterinarians. Devils with DFTD were obtained from the “Forestier trial” during which diseased devils were removed from the wild and euthanized through the program undertaken by the DPIW.
Differential Leukocyte Counts
Differential leukocyte counts were performed by the Pathology Department, Royal Hobart Hospital, Tasmania, Australia, using an automated counter (Technicon H2 Automated Counter, Dublin, Ireland).
Autopsies and Histology
Autopsies were performed on recent road kill carcasses that had been transported to the laboratory as rapidly as possible and immediately necropsied. The lymphoid tissues were removed and fixed in 10% neutral buffered formalin and paraffin embedded using the routine procedures of the Pathology Department, Royal Hobart Hospital, Tasmania Australia, and 3-μM sections were prepared and stained with hematoxylin and eosin and examined under light microscopy.
Mitogen-induced Lymphocyte Proliferation
Mononuclear cells were isolated from whole blood using a density gradient (Histopaque – 1077®; Sigma Chemical Co., St. Louis, MO) and centrifuged at 400g for 30 minutes. The mononuclear cell interface was washed twice in incomplete medium (RPMI 1640; JRH Biosciences, Inc., Lenexa, KS) containing 100 IU/ml gentamicin and 2 mM glutamine and diluted to 106 cells/ml. Autologous plasma (10%) was used as support for cell growth in all cultures. One hundred microliters of these cells was cultured in 96–U-bottomed-well plates (3870; Iwaki, Asahi, Japan) and incubated for 96 hours with 100 μg/ml phytohemagglutinin (PHA, L1668; Sigma Chemical Co., St. Louis, MO), 50 μg/ml concanavalin A (Con A, C7275; Sigma Chemical Co.), or 2 μg/ml pokeweed mitogen (PWM, L9379; Sigma Chemical Co.) and pulsed during the last 18 hours with 1 μCi [methyl-3H] thymidine (TR310; Amersham Pharmacia, Biotech, UK). Cells were then harvested onto a filter paper on a cell harvester (Skatron Combicell Harvester, Suffolk, UK), and tritiated thymidine incorporation as a measure of cell proliferation was determined with a 1214 Rackbeta liquid scintillation counter (LKB Wallac, Turku, Finland). Results are expressed as stimulation index (SI), where SI is (counts per minute stimulated cells) per (counts per minute unstimulated cells). All cultures were performed in triplicate.
Mixed Lymphocyte Reactions
Two-way mixed lymphocyte reactions were performed by preparing mononuclear cells (isolated as above) from the peripheral blood of captive devils into RPMI 1640 culture medium containing 100 IU/ml gentamicin and 2 mM glutamine and supplemented with 10% plasma, which was pooled from ten healthy devils. Cell concentration was adjusted to 106 cells/ml. One hundred microliters from two unrelated devils was added to 96–U-bottomed-well plates and incubated for 96 hours and pulsed during the last 18 hours with 1 μCi [methyl-3H] thymidine. Cells were then harvested onto filter paper and tritiated thymidine incorporation as a measure of cell proliferation was determined with a 1214 Rackbeta liquid scintillation counter. Results are expressed as stimulation index as above. All cultures were performed in triplicate.
Immune Response to Red Blood Cells
One male and one female Tasmanian devil received 0.5 ml of washed and packed horse red blood cells (HRBC) (defibrinated horse blood, Oxoid, Australia) plus 0.5 ml of Montanide ISA 775 VG (manufactured by Seppic, France, and provided by Tall Bennett Group) via the subcutaneous route at four sites on the back of the animal. A booster was administered 28 days later and the adjuvant was Montanide ISA 763A VG (manufactured by Seppic, France, and provided by Tall Bennett Group). Blood was collected for serum on a weekly basis for 8 weeks.
Hemagglutination Assay
Hemagglutination assays were performed in 96-V-bottomed-well plates (651101, Greiner Bio-One, Frankfurt, Germany) by adding serial dilutions of 25 μl immune devil serum to 25 μl of a solution of 2% washed HRBC and incubated at 37°C for 60 minutes. For indirect hemagglutination assays, 10 μl of a 1:20 dilution of rabbit anti-devil immunoglobulin (prepared in-house by immunizing rabbits with devil immunoglobulin) was added after the first 30 minutes of incubation. The titer was read as the highest antibody dilution that caused 50% agglutination. All assays were performed simultaneously.
Results
Immune Cells and Organs
The most accessible source for cells of the immune system is peripheral blood. Analysis of blood films demonstrated a predicted range of white blood cells including neutrophils, lymphocytes, monocytes, basophils, and eosinophils (Table 1). Postmortem examination of eight healthy disease-free devils revealed primary and secondary lymphoid organs. Thymus tissue was found adjacent to the heart and was clearly present in juvenile and subadult animals. Their lymphoreticular system comprised a spleen and numerous lymph nodes, providing anatomical support for a potentially functional immune system. A number of interesting observations were noted in the histology of these organs. Importantly, the histologic appearance of the spleen and lymph nodes is not dissimilar from placental mammals. The spleen contained classical areas of red and white pulp, and within the white pulp B-lymphocyte follicular activity and periarteriolar T-lymphocyte areas could be identified (Fig. 1). Lymph node assessment located B-cell follicles and cellular T cell areas within the cortex and paracortex respectively (Fig. 2). In both the spleen and lymph nodes, the B-cell areas appeared to be activated. In particular, lymph nodes were identified as possessing numerous secondary follicles and active germinal centers indicative of an active immune response. When 18 devils with DFTD were analyzed, a conspicuous feature of their lymphoid tissues was the presence of numerous plasma cells in lymph nodes, the spleen (Fig. 3), and occasionally the bone marrow.

Hematoxylin and eosin-stained histologic section of Tasmanian devil spleen showing an arteriole with periarteriolar lymphoid sheath and associated follicle. Scale bar, bottom left corner = 20 μM.

Hematoxylin and eosin-stained histologic section of Tasmanian devil lymph node showing cortex with numerous distinct B-cell follicles. Scale bar, top right corner = 100 μM.

Hematoxylin and eosin-stained histologic section of spleen from a diseased Tasmanian devil showing numerous plasma cell-like cells. Scale bar, bottom left corner = 20 μM.
Lymphocyte Proliferation
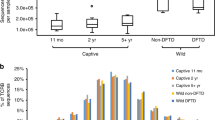
To investigate the specific immune response, lymphocyte function was analyzed in vitro by stimulating the proliferation of peripheral blood mononuclear cells (including lymphocytes) with mitogens, which are chemicals that bind to lymphocytes to induce cell division. The T-cell mitogens phytohemagglutinin (PHA) and concanavalin A (Con A) and the T- and B-cell mitogen pokeweed mitogen (PWM) all induced lymphocyte proliferation (Fig. 4). Thus, both T and B lymphocytes can be induced to proliferate in vitro and therefore appear to be functionally competent. However, when mixed lymphocyte reactions were performed, as an in vitro measure of an allogeneic T-cell response, there was a failure of devil lymphocytes to proliferate. As shown in Figure 4, the large standard deviations indicate a wide range of mitogen-induced lymphocyte proliferation among the devils. Hence, lymphocytes from different devils respond to different extents, indicating a diversity of responses among the devil population. From this information it is clear that lymphocytes isolated from the peripheral blood of Tasmanian devils have the capacity to proliferate when stimulated by mitogens, but not allogeneic lymphocytes.

Mitogen and two-way mixed-lymphocyte proliferation assays. Results represent mean ± standard deviation. Con A provided the greatest stimulation and the mean ± 1 standard deviation ranged from SI = 2 to SI = 349, whereas the mixed-lymphocyte reaction provided the weakest stimulation and the mean ± 1 standard deviation ranged from SI =1.0 to SI = 3.8. n = number of animals tested. Numbers in boxes represent the mean stimulation index for each group.
Antibody Response to Red Blood Cells
Assays of lymphocyte functions in vitro provide useful assessment of cell function, but this may not represent immune function in vivo. An effective measure of an animal’s immune function is to immunize the animal and evaluate the resultant antibody response. This was performed with the Tasmanian devil by subcutaneously injecting HRBCs in the presence of adjuvant and using hemagglutination to analyze the serum for the presence of specific antibodies. The subcutaneous immunization with HRBCs clearly demonstrated that male and female Tasmanian devils have the ability to produce a strong antibody response (Fig. 5). A booster injection at 28 days was found to increase the titer of antibodies.

Hemagglutination titers representing antibody production from a male and a female Tasmanian devil immunized subcutaneously with horse red blood cells in adjuvant at days 0 and 28 (indicated by arrows). Direct represents antibodies that agglutinate red cells on their own (IgM-like), whereas indirect represents the presence of all antibodies (IgM-like and IgG-like). Results show a rapid rise and a very high titer representing high levels of antibody production. Following the booster dose (day 28) there is evidence for a further increase in the total levels of antibody. Both the male (squares) and female devil (triangle) produced similar levels of antibodies in both the direct (open squares and open triangles) and the indirect (solid squares and solid triangles) hemagglutination assays.
Discussion
A potential explanation for devil-to-devil transfer of DFTD is that Tasmanian devils have a severely impaired immune system; however, this is unlikely as our investigations indicate a competent immune response. Examination of peripheral blood revealed an expected range of leukocyte populations, and autopsies performed on healthy devils as well as histology on normal primary and secondary lymphoid tissue clearly indicated that Tasmanian devils have all the necessary immune system components. Their lymphoid compartments are similar to those already described for other marsupials (Canfield and Hemsley, 2000). The histologic analysis of spleen and lymph nodes identified areas of T- and B-cell activity supporting active lymphocyte stimulation, particularly B cells. These observations suggest that DFTD occurs against a background of hyperactive B cells, and it remains unknown whether this B-cell response contributes to or is coincident with the malignant process. Further investigations are required to determine whether DFTD has a systemic immunopathology with a background of hyperactive B cells in addition to the localized tumor pathology observed in the facial area.
We have previously analyzed aspects of the innate immune response by evaluating phagocytosis of bacteria by peripheral blood neutrophils. This revealed a fully competent phagocytic system because neutrophils could bind, internalize, and phagocytose bacteria (Kreiss et al., unpublished). This is not surprising as devils are likely to be exposed to a variety of bacteria while they consume entire carcasses, including the entrails, which are colonized by a range of bacteria. Because phagocytosis is an important protective mechanism against bacteria, a healthy phagocytic system would be anticipated.
The histopathologic observations demonstrate that while devils have the necessary immunologic armor in the form of immune cells and tissues, there is rarely any lymphocyte infiltration into DFTD tumors. On the basis of CD3 expression to identify T cells, Loh and colleagues used immunohistochemistry on 18 devils and did not identify any T cells within any tumors (Loh et al., 2006b). This lack of T-lymphocyte infiltration, combined with the remarkable observation that DFTD is transferable, suggests that T-lymphocyte function may be impaired in these animals because although these cells are present, they may be unable to react appropriately. With a lack of specific reagents available to undertake a comprehensive analysis of the immune system, in vitro studies of immune function through mitogen-induced proliferation were initially used to investigate the specific immune response.
Healthy DFTD-free devils responded to the commonly used mitogens Con A, PHA, and PWM. The average stimulation indices were found to be similar to those reported for other eutherian mammals, including human, mouse, horse, red kangaroo, and tree kangaroo (Montali et al., 1998) as well as for koalas (Wilkinson et al., 1992), but significantly higher than that of the Tammar wallaby, although these cultures were undertaken in serum-free medium (Young and Deane, 2007). Consequently, lymphocytes isolated from the blood of Tasmanian devils are functionally competent as they have the ability to respond to mitogen stimulation. There was, however, an extremely diverse range of responses, with the lymphocytes from some animals responding very poorly. In separate work we found that when blood from these “poor responders” was collected again at a later date, the lymphocyte proliferative responses had improved substantially (Kreiss et al., unpublished). Therefore, a wide range of responses occurs with this species and within individual animals.
To evaluate an in vivo immune response to a cellular antigen, horse red blood cells were injected subcutaneously into a healthy male and a healthy female Tasmanian devil. Both devils produced high antibody titers, indicative of an effective antibody response, and following a second immunization at 28 days, the antibody levels were boosted. When these antibody levels are compared with that of another marsupial (the gray short-tailed opossum, Monodelphis domestica), the antibody response by the Tasmanian devil appears far superior as the antibody titers with the Monodelphis peaked at around 500 (Croix et al., 1989), whereas the antibody levels produced by the devil reached a maximum level of around 50,000, i.e., 100 times more effective.
Armed with this evidence that the devil can respond to a cellular antigen, the next step was to determine why DFTD fails to activate the devil’s immune system. To understand this, mixed lymphocyte reactions were performed. The mixed lymphocyte reaction is an in vitro measure of allogeneic responses that occur when a tissue transplant, or foreign tumor, is rejected. Devil lymphocytes failed to react to each other as the stimulation indices were approximately 2, which was at least 100-fold less than Con A stimulation. A poor mixed-lymphocyte reaction has been observed with the marsupial Monodelphis domestica (Stone et al., 1998), but Montali and colleagues undertook allogeneic mixed-lymphocyte reactions with a range of eutherian mammals and obtained stimulation indices similar to those they obtained following Con A stimulation (Montali et al., 1998). It is possible that poor allogeneic lymphocyte responses are characteristic of marsupials and the transmissible nature of DFTD could be a consequence of the poor allogeneic lymphocyte response, i.e., due to a failure of DFTD to be recognized by the devil’s immune system.
Allogeneic responses, as most immune responses, rely on the presence of a major histocompatibility complex (MHC), a multigene complex that codes for the ligands expressed on a range of cells to present antigen fragments to T cells (Belov et al., 2006). An absence of foreign MHC molecules would prevent the activation of T-cell responses to tumors and/or transplanted tissues. In many human cancers a downregulation of MHC Class Ia expression by the tumor cells allows these tumors to develop unhindered by the immune system (Algarra et al., 2000). A lack of MHC expression is unlikely to account for the failure of the devil’s immune system to reject the DFTD allografts because the tumor cells, which were analyzed by constructing a cDNA library, all expressed MHC class Ia and Class II genes (Siddle et al., unpublished). An alternative explanation is that there is a lack of genetic diversity within the devil population and the “cancer graft” MHC types are identical to those of the host. Work undertaken by Jones et al. (2004) supports the concept that devils have a low genetic diversity similar to cheetahs, one of the world’s most inbred species (Menotti-Raymond and O’Brien, 1995). In preliminary analyses of MHC class I genes, very little diversity was evident between different devils (Siddle et al., unpublished). Consequently, the best hypothesis to date that explains how DFTD can be transmitted between devils as an infectious allograft is a lack of diversity at MHC genes resulting in a failure of the DFTD tissue to be recognized as “non-self” by the host’s immune system.
The evidence is now overwhelming that DFTD is an infectious cancer and that the devil population is succumbing to a “tumor epidemic” of epic proportions. To control this epidemic, lessons can be learned from past epidemics in human populations and agricultural animals. Although analogies with other contagious diseases provide predictive models for DFTD, some unique considerations provide pertinent differences. The most important difference is the absence of convincing evidence of animals that are naturally resistant to this fatal tumor, hence a natural process for genetic selection is highly improbable. This separates DFTD from most other forms of infectious disease for which some level of resistance is present.
The biting and cannibalistic behavior of devils is most likely a major contributing factor for devil-to-devil transmission and clearly these wild animals cannot be re-educated to reduce contact rates. A small-scale trial is currently underway on a virtually isolated peninsula to remove diseased animals to break transmission of the disease, but to undertake such a program throughout the entire island of Tasmania has substantial logistical and financial implications. Rather than relying on removing the disease from the population, an alternative control measure would be to introduce resistance, either genetically or immunologically. The introduction of artificial immunity by means of vaccination is an alternative approach since vaccination remains a cornerstone of numerous protective health measures and its implementation has been significant with the protection against a diverse range of infectious diseases (Ada, 2005), both in human and animal populations. Because we consider DFTD within the realm of an infectious cancer (i.e., an infectious disease), vaccination needs to be considered an important protective strategy.
Although vaccination against diseases such as cancer has met with limited success, numerous experiments performed in mice have shown that tumors can be prevented in a well-controlled setting. For an effective vaccine many questions must be answered, including the identification of tumor antigens and whether the host can produce an effective antitumor T-cell response. Even when the host can produce effective antitumor immunity, numerous mechanisms remain that will allow tumor cells to escape immunologic destruction. These include the immune response with factors such as a lack of T-cell help, inadequate T-cell stimulation, presence of regulatory T cells, and failure of T cells to enter the tumor. Factors relating directly to the tumor include inadequate tumor antigen expression, lack of MHC expression, and production of immunosuppressive factors (Rosenberg, 2004). Consequently, the task of producing an effective vaccine is monumental; nonetheless, lessons learned from animal studies and human trials can assist in designing vaccine strategies.
There is evidence for and against vaccine and these points are summarized in Table 2. In support of vaccination as an effective strategy is the devil’s immune response itself. There is good evidence that the devil has an effective immune response, although it has yet to be determined whether cytotoxic T-cell activity can be induced. Further support for vaccination comes from the tumor which, as shown above, expresses MHC molecules which could be harnessed by the immune response to present antigen to T cells. However, the most encouraging aspect is that the tumor is highly conserved morphologically (Loh et al., 2006a) and genetically (Pearse and Swift, 2006). The transmissible nature and stability of the genome means that tumor antigens will be conserved and the immune response will have a constant antigenic target to react against, a critical factor for a successful vaccine approach.
The evidence against a successful vaccine approach lies within the restricted genetic diversity or, more specifically, the limited MHC diversity, and therefore the tumor and any associated tumor antigens are not recognized. Consequently, the immune system is not activated and the “transplanted” tumor is not rejected. This lack of MHC diversity was reflected in the failure to induce allogeneic lymphocyte proliferation in vitro, and without such recognition the ability to induce a response against this tumor is limited. The task ahead for a vaccine approach is to “trick” the devil’s immune system into recognizing the tumor as “non-self.” Even with a successful vaccine, an even more daunting challenge would be delivery of the vaccine in the wild so that a sufficient number of animals are vaccinated to induce “herd immunity,” thereby limiting the spread of the disease and protecting the nonimmune animals. Any vaccination strategy would need to be aimed at protecting accessible devil populations rather than eliminating the disease (Haydon et al., 2006).
References
Ada G (2005) Overview of vaccines and vaccination. Molecular Biotechnology 29:255–272
Algarra I, Cabrera T, Garrido F (2000) The HLA crossroad in tumor immunology. Human Immunology 61:65–73
Belov K, Deakin JE, Papenfuss AT, Baker ML, Melman SD, Siddle HV, et al. (2006) Reconstructing an ancestral mammalian immune supercomplex from a marsupial major histocompatibility complex. PLoS Biology 4:e46; DOI:10.1371/journal.pbio.0040046
Canfield PJ, Hemsley S (2000) The roles of histology and immunohistology in the investigation of marsupial disease and normal lymphoid tissue. Developmental and Comparative Immunology 24:455–471
Croix DA, Samples NK, Vandeberg JL, Stone WH (1989) Immune response of a marsupial (Monodelphis domestica) to sheep red blood cells. Developmental and Comparative Immunology 13:73–78
Das U, Das AK (2000) Review of canine transmissible venereal sarcoma. Veterinary Research Communications 24:545–556
Haydon DT, Randall DA, Matthews L, Knobel DL, Tallents LA, Gravenor MB, Williams SD, Pollinger JP, Cleaveland S, Woolhouse ME, Sillero-Zubiri C, Marino J, Macdonald DW, Laurenson MK (2006) Low-coverage vaccination strategies for the conservation of endangered species. Nature 443:692–695
Jones ME, Paetkau D, Geffen E, Moritz C (2004) Genetic diversity and population structure of Tasmanian devils, the largest marsupial carnivore. Molecular Ecology 13:2197–2209
Loh R, Bergfeld J, Hayes D, O’Hara A, Pyecroft S, Raidal S, Sharpe R (2006a) The pathology of devil facial tumor disease (DFTD) in Tasmanian Devils (Sarcophilus harrisii). Veterinary Pathology 43:890–895
Loh R, Hayes D, Mahjoor A, O’Hara A, Pyecroft S, Raidal S (2006b) The immunohistochemical characterization of devil facial tumor disease (DFTD) in the Tasmanian Devil (Sarcophilus harrisii). Veterinary Pathology 43:896–903
Menotti-Raymond MA, O’Brien SJ (1995) Evolutionary conservation of ten microsatellite loci in four species of Felidae. Journal of Heredity 86:319–322
Montali RJ, Bush M, Cromie R, Holland SM, Maslow JN, Worley M, Witebsky FG, Phillips TM (1998) Primary Mycobacterium avium complex infections correlate with lowered cellular immune reactivity in Matschie’s tree kangaroos (Dendrolagus matschiei). Journal of Infectious Diseases 178:1719–1725
Murgia C, Pritchard JK, Kim SY, Fassati A, Weiss RA (2006) Clonal origin and evolution of a transmissible cancer. Cell 126:477–487
Pearse AM, Swift K (2006) Allograft theory: transmission of devil facial-tumour disease. Nature 439:549
Rosenberg SA (2004) Development of effective immunotherapy for the treatment of patients with cancer. Journal of the American College of Surgeons 198:685–696
Stone WH, Brunn DA, Foster EB, Manis GS, Hoffman ES, Saphire DG, VandenBerg JL, Infante AJ (1998) Absence of a significant mixed lymphocyte reaction in a marsupial (Monodelphis domestica). Laboratory Animal Science 48:184–189
Wilkinson R, Kotlarski I, Barton M, Phillips P (1992) Isolation of koala lymphoid cells and their in vitro responses to mitogens. Veterinary Immunology and Immunopathology 31:21–33
Young LJ, Deane EM (2007) Culture and stimulation of tammar wallaby lymphocytes. Veterinary Research Communications 31:685–701
Acknowledgments
The authors gratefully acknowledge all members of the Devil Facial Tumour Disease Team. They also thank Dr. Barrie Wells and Dr. James Harris for veterinary assistance and Candice Clark for technical assistance. They also acknowledge the assistance provided by the Pathology Department, Royal Hobart Hospital. Financial support for this work came from the Department of Primary Industry and Water and research grants from the Eric Guiler Foundation, the Australian Research Council, and the Sarah and Anne Payten Canine Cancer Fund. Ethics was approved by the Animal Ethics Committee of Tasmanian’s Park and Wildlife Services, permit numbers 33/2004–5 and 32/2005–6
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Woods, G.M., Kreiss, A., Belov, K. et al. The Immune Response of the Tasmanian Devil (Sarcophilus harrisii) and Devil Facial Tumour Disease. EcoHealth 4, 338–345 (2007). https://doi.org/10.1007/s10393-007-0117-1
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10393-007-0117-1